|
|
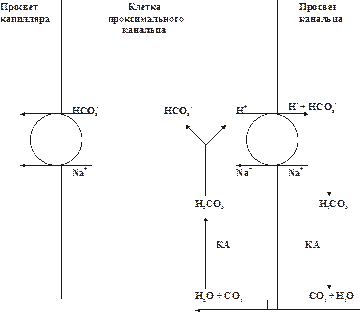
Гемоглобиновая буферная системаГемоглобиновая буферная система — наиболее ёмкий буфер крови — составляет более половины всей её буферной ёмкости. Гемоглобиновый буфер состоит из кислого компонента — оксигенированного Hb — HbO2 и основного — неоксигенированного. HbO2 примерно в 80 раз сильнее диссоциирует с отдачей в среду H+, чем Hb. Соответственно, он больше связывает катионов, главным образом K+. Основная роль гемоглобиновой буферной системы заключается в её участии в транспорте CO2 от тканей к лёгким. В капиллярах большого круга кровообращения HbO2 отдаёт кислород. В эритроцитах CO2 взаимодействует с H2O и образуется H2CO3. Эта кислота диссоциирует на HCO3– и H+, который соединяется с Hb. Анионы HCO3– из эритроцитов выходят в плазму крови, а в эритроциты поступает эквивалентное количество анионов Cl–. Остающиеся в плазме крови ионы Na+ взаимодействуют с HCO 3– и благодаря этому восстанавливают её щелочной резерв. В капиллярах лёгких, в условиях низкого pСО2 и высокого pО2, Hb присоединяет кислород с образованием HbO2. Карбаминовая связь разрывается, в связи с чем высвобождается CO2. При этом, HCO3– из плазмы крови поступает в эритроциты (в обмен на ионы Cl–) и взаимодействует с H+, отщепившимся от Hb в момент его оксигенации. Образующаяся H2CO3 под влиянием карбоангидразы расщепляется на CO2 и H2O. CO2 диффундирует в альвеолы и выводится из организма. Карбонаты костной ткани Карбонаты костной ткани функционируют как депо для буферных систем организма. В костях содержится большое количество солей угольной кислоты: карбонаты кальция, натрия, калия и др. При остром увеличении содержания кислот (например, при острой сердечной, дыхательной или почечной недостаточности, шоке, коме и других состояниях) кости могут обеспечивать до 30–40% буферной ёмкости. Высвобождение карбоната кальция в плазму крови способствует эффективной нейтрализации избытка H+. В условиях хронической нагрузки кислыми соединениями (например, при хронической сердечной, печёночной, почечной, дыхательной недостаточности) кости могут обеспечивать до 50% буферной ёмкости биологических жидкостей организма. ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ Наряду с мощными и быстродействующими химическими системами в организме функционируют органные механизмы компенсации и устранения сдвигов КОС. Для их реализации и достижения необходимого эффекта требуется больше времени — от нескольких минут до нескольких часов. К наиболее эффективным физиологическим механизмам регуляции КОС относят процессы, протекающие в лёгких, почках, печени и ЖКТ. Лёгкие Лёгкие обеспечивают устранение или уменьшение сдвигов КОС путём изменения объёма альвеолярной вентиляции. Это достаточно мобильный механизм — уже через 1–2 мин после изменения объёма альвеолярной вентиляции компенсируются или устраняются сдвиги КОС. Причиной изменения объёма дыхания является прямое или рефлекторное изменение возбудимости нейронов дыхательного центра. Снижение рН в жидкостях организма (плазма крови, СМЖ) является специфическим рефлекторным стимулом увеличения частоты и глубины дыхательных движений. Вследствие этого лёгкие выделяют избыток CO2 (образующийся при диссоциации угольной кислоты). В результате содержание H+ (HCO3– + H+ = H2CO3 Повышение рН в жидких средах организма снижает возбудимость инспираторных нейронов дыхательного центра. Это приводит к уменьшению альвеолярной вентиляции и выведению из организма CO2, т.е. к гиперкапнии. В связи с этим в жидких средах организма возрастает уровень угольной кислоты, диссоциирующей с образованием H+, — показатель рН снижается. Следовательно, система внешнего дыхания довольно быстро (в течение нескольких минут) способна устранить или уменьшить сдвиги рН и предотвратить развитие ацидоза или алкалоза: увеличение вентиляции лёгких в два раза повышает рН крови примерно на 0,2; снижение вентиляции на 25% может уменьшить рН на 0,3‑0,4. Почки К главным механизмам уменьшения или устранения сдвигов КОС крови, реализуемых нефронами почек, относят ацидогенез, аммониогенез, секрецию фосфатов и K+,Na+‑обменный механизм. • Ацидогенез. Этот энергозависимый процесс, протекающий в эпителии дистальных отделов нефрона и собирательных трубочек, обеспечивает секрецию в просвет канальцев H+ в обмен на реабсорбируемый Na+ (рис. 14–1). Ы ВЁРСТКА Вставить файл «ПФ Рис 14 01 Реабсорбция HCO3‑ в клетках проксимального отдела»
Рис.14–1.Реабсорбция HCO3‑ в клетках проксимального отдела. КА — карбоангидраза. Ы ВЁРСТКА Вставить файл «ПФ Рис 14 02 Реабсорбция HCO3‑ в клетках проксимального отдела»
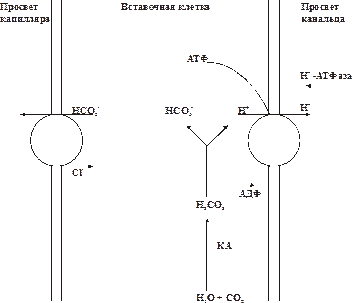
Рис.14–2.Секреция H+ клетками канальцев и собирательных трубочек. КА — карбоангидраза. Количество секретируемого H+ эквивалентно его количеству, попадающему в кровь с нелетучими кислотами и H2CO3. Реабсорбированный из просвета канальцев в плазму крови Na+ участвует в регенерации плазменной гидрокарбонатной буферной системы (рис. 13–2). • Аммониогенез, как и ацидогенез, реализует эпителий канальцев нефрона и собирательных трубочек. Аммониогенез осуществляется путём окислительного дезаминирования аминокислот, преимущественно (примерно 2/3) — глютаминовой, в меньшей мере — аланина, аспарагина, лейцина, гистидина. Образующийся при этом аммиак диффундирует в просвет канальцев. Там NH3+ присоединяет ион H+ с образованием иона аммония (NH4+). Ионы NH4+ замещают Na+ в солях и выделяются преимущественно в виде NH4Cl и (NH4)2SO4. В кровь при этом поступает эквивалентное количество гидрокарбоната натрия, обеспечивающего регенерацию гидрокарбонатной буферной системы. • Секреция фосфатов осуществляется эпителием дистальных канальцев при участии фосфатной буферной системы: Na2HPO4 + H2CO3 Образующийся гидрокарбонат натрия реабсорбируется в кровь и поддерживает гидрокарбонатный буфер, а NaH2PO4 выводится из организма с мочой. Таким образом, секреция H+ эпителием канальцев при реализации трёх описанных выше механизмов (ацидогенеза, аммониогенеза, секреции фосфатов) сопряжена с образованием гидрокарбоната и поступлением его в плазму крови. Это обеспечивает постоянное поддержание одной из наиболее важных, ёмких и мобильных буферных систем — гидрокарбонатной и как следствие — эффективное устранение или уменьшение опасных для организма сдвигов КОС. • К+,Na+‑обменный механизм, реализуемый в дистальных отделах нефрона и начальных участках собирательных трубочек, обеспечивает обмен Na+ первичной мочи на K+, выводящийся в неё эпителиальными клетками. Реабсорбированный Na+ в жидких средах организма участвует в регенерации гидрокарбонатной буферной системы. K+,Na+‑обмен контролируется альдостероном. Кроме того, альдостерон регулирует (увеличивает) объём секреции и экскреции H+. Таким образом, почечные механизмы устранения или уменьшения сдвигов КОС осуществляются путём экскреции H+ и восстановления резерва гидрокарбонатной буферной системы в жидких средах организма. Печень Печень играет существенную роль в компенсации сдвигов КОС. В ней действуют, с одной стороны, общие внутри‑ и внеклеточные буферные системы (гидрокарбонатная, белковая и др.), с другой стороны, в гепатоцитах осуществляются различные реакции метаболизма, имеющие прямое отношение к устранению расстройств КОС. • Синтез белков крови, входящих в белковую буферную систему. В печени образуются все альбумины, а также фибриноген, протромбин, проконвертин, проакцелерин, гепарин, ряд глобулинов и ферментов. • Образование аммиака, способного нейтрализовать кислоты как в самих гепатоцитах, так и в плазме крови и в межклеточной жидкости. • Синтез глюкозыиз неуглеводных веществ — аминокислот, глицерина, лактата, пирувата. Включение этих органических нелетучих кислот при образовании глюкозы обеспечивает снижение их содержания в клетках и биологических жидкостях. Так, МК, которую многие органы и ткани не способны метаболизировать, в гепатоцитах примерно на 80% трансформируется в H2O и CO2, а оставшееся количество ресинтезируется в глюкозу. Таким образом, лактат превращается в нейтральные продукты. • Выведение из организма нелетучих кислот — глюкуроновой и серной при детоксикации продуктов метаболизма и ксенобиотиков. • Экскреция в кишечниккислых и основных веществ с жёлчью. Желудок и кишечник Желудок участвует в демпфировании сдвигов КЩР, главным образом, путём изменения секреции соляной кислоты: при защелачивании жидких сред организма этот процесс тормозится, а при закислении — усиливается. Кишечник способствует уменьшению или устранению сдвигов КЩР посредством: • Секреции кишечного сока, содержащего большое количество гидрокарбоната. При этом в плазму крови поступает H+. • Изменения количества всасываемой жидкости. Это способствует нормализации водного и электролитного баланса в клетках, во внеклеточной и других биологических жидкостях и как следствие — — нормализации рН. • Реабсорбции компонентов буферных систем (Na+, K+, Ca2+, Cl–, HCO3–). 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|