|
|
БИОФИЗИЧЕСКИЙ АСПЕКТ НОМОГЕНЕЗАК настоящему времени в современной биологии накопилось достаточно фактов о процессах видообразования, о них создано много модельных представлений, которые свидетельствуют о том, что ни одна из существующих гипотез эволюции живого не может дать исчерпывающий ответ на все механизмы возникновения и организации живого на Земле. Некоторые биологи считают, что на сегодняшний день уже нельзя противопоставлять имеющиеся гипотезы эволюции. Каждый из механизмов, описываемых той или иной гипотезой, объясняет один или группу процессов, происходящих в реальности. Современное понимание взаимоотношений между тремя наиболее известными учениями таково представлено на рис. П 1. Исследования количественных характеристик признаков видов, родов, отрядов, семейств показали, что «родственные» группы объединяются друг с другом в образования, имеющие основное плотное ядро и диффузную периферию, плотность заполнения которой весьма мала. Такие распределения обычно описываются законом плотности распределения Паретто. Это наводит исследо-
вателей на мысль, что новые группы возникают скачком, образующим основное ядро, а затем происходит заполнение периферии по определенным правилам. СВ. Мейен, исследуя проблему разнообразия таких параллельных рядов, показал их аналогию с грамматиками различных языков. Имеются корни слов (основные ядра, образующие группы устойчивых признаков каждого вида), которые в различных падежах не изменяются, а падежные окончания (периферия основных ядер) однотипно и закономерно сменяют друг друга, несмотря на различия в ядрах. Этот феномен был назван им «биологической грамматикой». Подобным образом можно строить таблицы, где, например, строки указывают вариации признака, а столбцы – виды, роды, отряды или семейства. Выяснение законов «биологической грамматики» едва начато. И многие исследователи считают, что саму систематику следует строить так, чтобы таксон (раса, подвид, вид, род и т.д.) задавался пересечением рядов (табл. П 1). Эволюция, воспроизводя разнообразие видов, как бы заполняет ячейки таких таблиц. Параллели в природе могут быть настолько неожиданными, что сознание современного человека с трудом воспринимает это и, чаще всего, относит их к случайностям. Например, Г.Г. Длясиным обнаружен параллелизм между рядами свойств звуков азбуки алфавитов большинства различных языков и аминокислотами, являющимися кирпичиками первичной структуры белков. Он приводит удивительные параллели структуры звукоряда и различных физико-химических свойств аминокислот.
Таблица П1 Морфологический межклассовый рефрен мерона
С.В. Мейеном предложено учение о меронах, мероно-мия, где основным является понятие об архетипе. Мерон (от греч. meros – часть) – это «класс частей». Комбинации сочетаний меронов складывают обобщенный образ животного (архетип), например, птица, зверь, насекомое, червь и т. п. Эволюция реализуется чаще всего возникновением нового столбца, но прогрессивная эволюция всегда включает появление новой строки (и связанных с нею новых столбцов). СВ. Мейен, рассматривая параллелизмы рядов, предложил учитывать не столько повторность самих признаков, сколько повторность «законов преобразования» этих признаков. Ссылаясь на «номогенез» Л.С. Берга, Заварзин утверждал, что формы, имеющие различное происхождение, «в своем развитии не могут беспорядочно варьировать, а эволюционируют в одном более или менее направлении, почему в различных параллельных рядах и развиваются конвергирующие признаки. Таким образом, с этой точки зрения даже многие гомологические части обязаны своим сходством конвергенции», на что, в частности, указывает закон Вавилова. Коэволюция.Коэволюционная концепция рассматривает эволюцию данного объекта (например, вида) как ячейку в общем процессе, и это, пожалуй, главное отличие от прежнего эволюционизма, сосредоточенного на эволюции отдельных видов, якобы изменяющихся в довольно безликой среде. Экологи говорят о коэволюции давно, но у них это понятие означает лишь взаимное приспособление видов и не повлекло концептуальных изменений, тогда как расширение смысла коэволюции Янчем отражает новое понимание эволюции. Ивановский, Шургин и Обут и многие другие указывают на то, что в ряде случаев перестройка живого вещества предваряет крупные геологические события или же приходится на очень раннюю их фазу. Причем существуют как несинхронность тектонических и палеонтологических рубежей, так и развитие крупных флористических и фаунических комплексов. В ряде случаев крупные перестройки флоры совершаются раньше крупных перестроек фауны. Примером может служить меловой период. Покрытосеменные появились в середине мела и к концу его заняли доминирующее положение. Вымирание динозавров и других форм приходится в основном на верхний мел, и с этого времени начинается бурная эволюция млекопитающих. Суша в девоне вначале была освоена растениями и лишь затем позвоночными. Все это указывает на опережающую реакцию живого на будущие изменения окружающей среды, включая геологические катаклизмы планетарного масштаба. В ряде случаев в активных фазах циклов перед крупными перестройками экосистем широко развивается гигантизм. Он представляет интерес как своеобразная альтернатива истинному ароморфозу, основанному на глубоких внутренних системных изменениях. Чаще всего сверхразвитые гиганты оказываются тупиковыми формами и вымирают. Глубокие ароморфозные изменения захватывают в основном мелкие формы. Развитие гигантизма у независимых групп можно трактовать как довод в пользу существования некоего общего активирующего фактора, предшествующего образованию новых типов и классов живого. А.Г. Теслинов, рассматривая онтологию развития идеи, пишет: «Следует различать адаптационный и бифуркационный характер процессов развития. С первым связывают постепенные приспособительные изменения объектов под воздействием развивающих факторов. Бифуркационный характер развития соответствует катастрофическому, революционному изменению качества как разветвления вариантов эволюции... В эволюционном движении любого естественного процесса один характер последовательно сменяется другим». Таким образом, эволюция должна рассматриваться в неразрывной связи с эволюцией косного вещества не только Земли, но всей Солнечной системы, включая адаптационные и бифуркационные явления в процессе коэволюции Вселенной в целом. Наиболее полно системную картину-схему развития живого предложил Ю.В. Чайковский (рис. П 2).
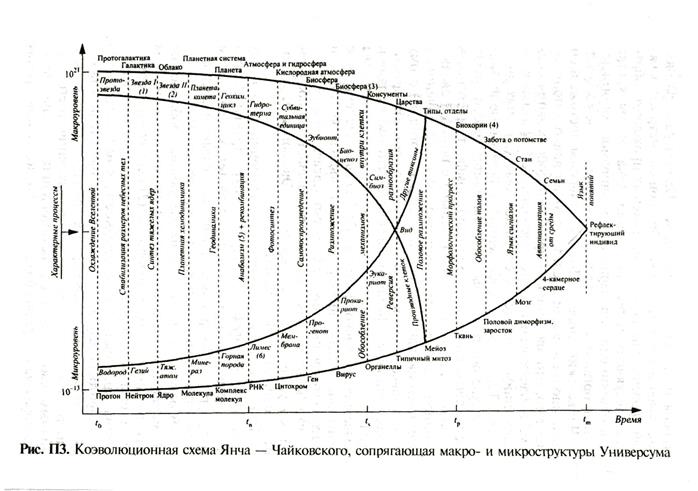
В его логической схеме диатропической четырехцарственной макросистемы организмов биосферы Земли отражены все наиболее существенные классификационные признаки и уровни усложняющейся оргашязации живого. Схема построена на кольцевой симметрии. Процедура образования колец тоже обладает своеобразной симметрией; каждый уровень, считая от одноклеточных прокариот, образует следующий путем интеграции: эукариотная клетка есть объединение прокариотных механизмов, многоклеточный организм или ткань – объединение клеток и т.д. Чайковский пишет: «Пожалуй, главным достижением эволюционной мысли последних 30 лет явилось понимание того, что эволюция – процесс преобразования систем, а наблюдаемый поток меняющихся признаков – лишь эпифеномен (сопутствующее явление) этого процесса... Теперь, когда собственно эволюционные физиологические понятия даны, вырисовывается совсем другая схема (эволюционного механизма): комфорт, инертность и сытость в равной степени гибельны с позиций эволюционной перспективы, стремление к неограниченному размножению наблюдается лишь как патология, а потому борьба вовсе не является основным фактором взаимодействия. Основным физиологическим фактором эволюции выступает дискомфорт. Существенно, что организмы подвергаются дискомфорту в тем большей степени, чем менее они приспособлены к своим условиям обитания, и в этом смысле был прав Э.С. Бауэр, писавший в 1935 году, что "материал для эволюции поставляют не победители (как у Дарвина), а побежденные... Эволюция не состоит из одних приспособлений"». Ю.В. Чайковский приводит коэволюционную схему (рис. П 3), сопрягающую макро- и микроструктуры, причем с течением времени в целом растет размер систем микроуровня (от фотонов до людей) и падает размер систем макроуровня (от протогалактик до семей). «В основу схемы положены следующие соображения: (1) метаболизм предшествовал наследованию и возник на базе геохимических круговоротов; (2) первичный метаболизм был гетеротрофным (потреблял небиологические потоки органики), но автотрофность (в том числе, вероятно, фотосинтез) возникла до начала экспансии живого, т.е. до появления самовоспроизведения; (3) генетический материал возник ранее самовоспроизведения, и его пер- f вичным свойством была рекомбинация, обеспечившая преобразование разнообразия еще до появления прочихфункций живого; (4) самовоспроизведение (размножение генетического материала) появилось до сегрегации живого на отдельные организмы; (5) значение ядра клетки сказалось, прежде всего, в симбиозе (прокариоты к симбиозу друг с другом неспособны), но затем оказалось главным в порождении многоклеточных (реверсия разнообразия).
Вся эволюция эукариот прошла между двумя системными рубежами. Момент (в геологическом смысле этого слова) ts характеризовался рождением симбиоза, а всякий симбиотический организм является микробиоценозом, поскольку между организмами, его составляющими, существуют круговороты. Следовательно, в момент ts объект микроуровня (организм) стал элементом макроуровня (биоценозом); после этого переплетение макро- и микроуровня нарастало; с появлением в момент tm человека деление эволюции на эти уровни просто потеряло смысл. Итак, эволюция эукариот, в основном законченная, отличалась от всех предыдущих переплетением уровней эволюционных процессов, а будущая эволюция (неизбежная, пока существует сама жизнь) должна описываться (если не исчезнет человечество) другим языком, о котором можно будет говорить только после выявления языка прежней эволюции». Астрофизический аспект номогенеза. Начиная с первых систематиков были попытки создавать общие системы, включающие косную и живую материю. Так, Генрих Брони, ботаник, зоолог и палеонтолог, первый критик и переводчик Дарвина в Германии, предложил систему «четырех царств природы» (небесных тел, минералов, растений и животных). Она была основана на «силах», действующих в этих царствах. Идея неслучайности возникновения и развития живого вдохновляла многих исследователей на осмысление общего принципа развития Универсума. Многие русские ученые (К.Э. Циолковский, В.И. Вернадский, А.Л. Чижевский) были убеждены, что проблемы эволюции биосферы необходимо рассматривать в контексте законов изменения Космоса. Открытие проблемы больших чисел и ее анализ показали, что существует жесткая привязанность всех параметров Вселенной к биологической жизни.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|