|
|
Регуляция активности геновКак могут существовать участки, гиперчувствительные к ДНКазе I, если ДНК единообразно упакована в нуклеосомы? На этот вопрос возможны два ответа. Первый в хроматине существуют области, свободные от нуклеосом, и второй – существуют факторы, которые присоединяются к нуклеосомам и заставляют их ослаблять связь с ДНК. Отсутствие нуклеосом обеспечивает доступность всей ДНК в этой области для РНК-полимеразы и возможность осуществления эффективной транс-
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. ___________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ___________________________ 165
крипции. У клопа-наземника Oncopeltus fasciatus часть генома, содержащая гены рибосомной РНК, практически полностью свободна от нуклеосом и ДНК в этой области транскрибируется очень активно (Foe et al., 1976). Результаты электронно-микроскопических исследований подтвердили это наблюдение для Oncopeltus и некоторых других видов (рис. 12.27) (Labhart, Koller, 1982). Транскрипция глобиновых генов мыши тоже может происходить с нарушениями в распределении
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 166_______________ ГЛАВА 12______________________________________________________________________________
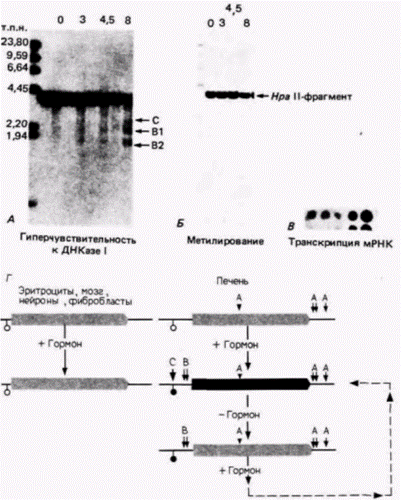
нуклеосом. В клетках, которые не относятся к эритроидному ряду, нуклеосомы на всем протяжении глобиновых генов располагаются регулярно. Однако в предшественниках эритроцитов регулярное расположение нуклеосом нарушается, так что 5'-конец гена (от -200 до +500) оказывается свободным от этих структур. Когда клетки индуцируются к синтезу гемоглобина, может происходить дальнейшее разрушение нуклеосом (Cohen, Sheffery, 1985; Benezra et al., 1986). Данные, полученные в других исследованиях, также свидетельствуют о том, что большинство участков, гиперчувствительных к ДНКазе I (а возможно, и все такие участки), лишены нуклеосом. Гены легких цепей иммуноглобулинов становятся гипометилированными и чувствительными к ДНКазе I в ходе развития пре-В-клеток в В-клетки (Rose, Garrard, 1984). Кроме того, в это время меняется распределение нуклеосом, так что большие областииммуноглобулинового гена становятся относительно свободными от них. Остающиеся нуклеосомы оказываются без гистона H1 и содержат вместо него белки HMG 14 и HMG 17 (см. ниже). Не исключено, что способ взаимодействия метильных групп с гистонами позволяет образовывать нуклеосомы только на метилированной ДНК, но не на неметилированной (Keshet et al., 1986). Это ограничение будет приводить к появлению участков гиперчувствительности к ДНКазе. После образования таких участков другим транс-регуляторным фактором станет проще находить эти свободные от нуклеосом области. Такая последовательность событий могла бы объяснить, почему ген вителлогенина в печени курицы содержит ряд сайтов, гиперчувствительных к ДНКазе, уже перед добавлением гормона. Присоединение транс-регуляторного рецептора гормона будет создавать впоследствии новые гиперчувствительные сайты, связанные с активной транскрипцией. Формирование активного участка хроматина не требует полного удаления нуклеосом. Многие активно транскрибируемые гены лежат внутри районов с обычным распределением нуклеосом. Тем не менее нуклеосомы могут быть модифицированы в определенных специфических локусах. Одна из таких наиболее важных модификаций включает присоединение двух пептидов, называемых белками группы высокой подвижности: HMG 14 и HMG 17. Когдахроматин гилролизуется ДНКазой I, происходит преимущественное высвобождение двух негистоновых белков хроматина: HMG 14 и HMG 17. Эти два низкомолекулярных белка могут быть экстрагированы из хроматина с помощью 0.35 M NaCl Они, по-видимому, обеспечивают чувствительность, но не гиперчувствительность генов к ДНКазе I, когда связываются поблизости от нуклеосом. Если
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. ____________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ___________________________ 167
белки H MG 14 и H MG 17 удаляют из хроматина, то одновременно исчезает дифференциальная чувствительность специфических генов к ДНКазе I. Таким образом, когда белки HMG экстрагированы из хроматина эритроцитов, глобиновые гены утрачивают чувствительность к перевариванию ДНКазой I (рис. 12.28). Однако, когда эти белки добавляют к такому обедненному хроматину, дифференциальная чувствительность глобиновых генов восстанавливается (Weisbrod, Weintraub, 1979; Weisbrod et al., 1980; Gazit et al., 1980). Способность этих белков восстанавливать чувствительность по-прежнему определяется хроматином. HMG 14 и HMG 17, выделенные из мозга, восстанавливают чувствительность к ДНКазе I глобиновых генов из эритроцитов, но не из клеток мозга. Таким образом, два негистоновых белка хроматина оказываются необходимыми (но не достаточными) для осуществления транскрипции специфических генов в разных тканях. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|