|
|
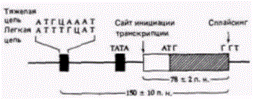
Транскрипция генов легких цепей иммуноглобулиновОдин из наиболее хорошо изученных случаев регуляции генов на уровне транскрипции относится к иммуноглобулиновым генам мыши. За последние пять лет были идентифицированы цис· и транс-регуляторные элементы, необходимые для специфической транскрипции этих генов в В-клетках. Делеционное картирование клонированных генов показало, что промотор легкой цепи иммуноглобулина имеет два критических участка – ТАТА-бокс и передний элемент, называемый «окта»-боксом (Bergman et al., 1984; Parslow et al., 1984). Окта-бокс, названный так потому, что состоит из 8 пар оснований, прочитывается как ΑТТТГЦAT. Он был обнаружен во всех изученных промоторах легких цепей иммуноглобулинов и в очень незначительном числе других сайтов. Промоторы генов тяжелых цепей иммуноглобулинов имеют «инвертированный» окта-бокс, АТГЦАААТ (рис. 12.15). Если окта-бокс помещают с 5'-стороны от глобинового гена, то транскрипция этого гена в клетке миеломы увеличивается в 11-18 раз. Это увеличение наблюдалось только в лимфоидных клетках и не наблюдалось в фибробластах (Wirth et al., 1987). Последовательность энхансера генов легких цепей иммуноглобулинов расположена внутри первого интрона между последовательностью VJ и последовательностью константной области (Queen, Baltimore, 1983; Bergman et al., 1984). Если эту последовательность транслировать на клонированные глобиновые гены, то эти гены также приобретают способность специфически транскрибироваться в лимфоцитах (Picard, Schaffner, 1984).
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 152_______________ ГЛАВА 12_____________________________________________________
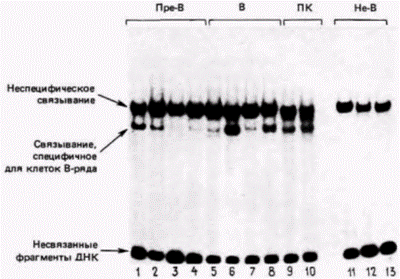
При созревании лимфоцитов в ходе перестроек генов происходит сближение энхансеров и промоторов. Количество ядерных транскриптов с перестроенных генов легких цепей (имеющих сближенные промотор и энхансер) более чем в 16 000 раз превышает количество транскриптов с генов, которые не перестроены (Mather, Perry, 1982). При этом энхансер легких цепей иммуноглобулинов стимулирует свои собственные промоторы в 20 раз эффективнее, чем промоторы вируса SV40 или гена металлотионеина. Это наблюдение предполагает синергизм в специфическом взаимодействии между обоими элементами (Garcia el al., 1986). Перестройки сами по себе еще не обеспечивают активацию генов, поскольку перестроенный ген иммуноглобулина не транскрибируется после введения его в фибробласт или в клетку печени. Для начала его транскрипции необходимо присутствие специфических для клетки транс-регуляторных факторов. В 1986 г. были идентифицированы два фактора, связывающиеся с промотором (Staudt et al., 1986). С целью идентификации небольшой фрагмент ДНК, содержащий окта-бокс, инкубировали в ядерных экстрактах из различных клеток. Затем полученные продукты фракционировали в геле. Если ядерный экстракт не содержал белка, связывающегося с этой ДНК, то небольшой фрагмент ДНК быстро двигался через гель. Однако если какой-либо белок действительно присоединялся к этой ДНК, то ее миграция через гель затруднялась. Этот опыт по сдвигу подвижности (рис. 12.16) показал, что каждое ядро содержит по крайней мере один фактор, способный присоединяться к данному фрагменту ДНК. Кроме того, клетки B-ряда (пре-В-клетки, В-клетки и плазматические клетки) дополнительно содержат еще один фактор, специфически связывающийся с ДНК окта-бокса. Эти ДНК-связывающие белки были изолированы, и белок, специфический
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ__________________________ 153
для лимфоцитов, был назван NF-A2. Аналогичные методы были использованы для поиска ядерного белка, характерного для клеток B-ряда и специфически связывающегося с энхансерными последовательностями генов легких цепей иммуноглобулинов (Sen, Baltimore, 1986a; Atchinson, Perry, 1987). Этот белок, обозначенный как NF-κΒ, обнаруживается только в зрелых B-клетках и плазматических клетках, т.е. в единственных известных клетках, синтезирующих иммуноглобулины 1. Активный NF-κΒ не обнаружен в клетках какого-либо другого типа (в том числе и в пре-В-клетках), а также в линии плазматических клеток, которые утратили способность синтезировать легкую цепь иммуноглобулинов. Активный белок NF-κΒ появляется тогда, когда пре-В-клетки индуцируются к образованию В-клеток. Эти наблюдения поднимают проблему дифференциальной активности генов на другой уровень: что именно контролирует синтез специфических для B-клеток белков – промоторсвязывающего белка NF-A2 и энхансерсвязывающего белка NF-ϰВ? (Иными словами, чтобы объяснить, как осуществляется специфический для клеток синтез иммуноглобулинов, мы должны теперь объяснить, как эти два ядерных белка синтезируются специфическим для клеток образом.) Оказалось, что белок NF-κΒ в неактивной форме содержится в клетках многих типов, но модифицироваться в активную форму он может только в B-клетках. Когда пре-В-клетки искусственно стимулируются к образованию В-клеток, активный NF-κΒ появляется даже в отсутствие белкового синтеза (Sen, Baltimore, 1986b). Природа модификации этого белка остается неизвестной. Анализ специфической для клеток транскрипции генов легкихцепей иммуноглобулинов вышел в итоге за уровень исследования продуктов терминальной дифференцировки клеток. Мы знаем, что транскрипция этого иммуноглобулинового гена зависит от исходной активности двух ядерных белков, NF-A2 и NF-κΒ, которая наблюдается только в клетках B-ряда. Остается выяснить, каким образом вызывается дифференциальная экспрессия этих двух генов в развивающемся лимфоците. Исследование иммуноглобулинового энхансера дало объяснение и другим биологическим явлениям. Ниже (т. 3. гл. 20) мы обсудим вопрос о том, что
1 В пре-В-клетке синтезируется только тяжелая цепь иммуноглобулинов и не синтезируется легкая. Вызывает удивление присутствие неспецифического белка, связывающегосяс окта-боксом, в клетках, не относящихся к В-ряду. Оказалось, что ген гистона Н2В имеет в своем энхансере окта-бокс, и присоединение неспецифическою белка, связывающегося с окта-боксом, определяет время его экспрессии, благодаря чему этот ген транскрибируется в течение S-фазы клеточного цикла (Roeder, 1987; Fletcher et al., 1987).
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 154_______________ ГЛАВА 12_____________________________________________________________________________
лимфоцитарные опухоли возникают, когда обычный ген фактора ростапереносится в область иммуноглобулиновогоэнхансера, благодаря чему ген фактора роста транскрибируется по времени и по количествукак антитела. Кроме того, быстрое размножение вируса СПИда, HIV-1, объясняется тем, что он имеет энхансер, очень похожий на энхансер легких цепей иммуноглобулинов. Когда этот вирус инфицирует Т-лимфоцит, он способен индуцировать образование активного NF-κΒ (Sen, Baltimore, 1986b). 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|