|
|
Дополнительные сведения и гипотезы: Определение сайтов метилированияКогда происходит метилирование ДНК, оно практически всегда идет по цитидину из ЦфГ-пары. Распределение этих метилированных и неметилированных ЦГ-дуплетов определяется с помощью разрезания ДНК двумя рестриктазами. HpaIIи MspI(McGhee, Ginder, 1979). Оба этих фермента разрезают в одном сайте, ЦЦГГ, но НраIIне будет разрезать ДНК, если метилирован центральный Ц, тогда как MspIбудет разрезать эту последовательность независимо от того, метилирована она или нет. Следовательно, ДНК из клеток определенного типа может быть гидролизована HpaIIи отдельно MspI;затем фрагменты расщепленной ДНК могут быть разделены с помощью электрофореза в геле, перенесены на нитроцеллюлозный фильтр и гибридизованы с радиоактивным зондом, специфичным для исследуемого гена. Разницу в распределении зон на радиоавтографах для фрагментов, полученных с MspIи с HpaII, можно объяснить разницей в метилировании. На рис. 12.21 показан результат одного из экспериментов с выделением ДНК спермиев и ее обработкой HpaIIи MspI. Зондом являлась радиоактивная ДНК из второго экзона ß-глобинового гена. Радиоавтограф фрагментов MspI-гидролизата
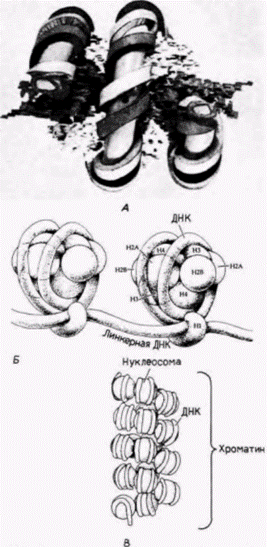
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ___________________________ 159 показывает, что этот зонд присоединяется к фрагментам ДНК, у которой ЦЦГГ-сайты лежат друг от друга на расстоянии 1400 пар оснований. Радиоавтограф с HpaII-гидролизатом показывает, что в ДНК спермиев эти сайты метилированы и что исследуемая последовательность ДНК лежит на отрезке ДНК длиной 25 000 пар оснований, на котором все ЦЦГГ-сайты метилированы (Groudine, Conklin, 1985). ХРОМАТИН ЭУКАРИОТ Нуклеосомы До сих пор при рассмотрении транскрипции мРНК мы ограничивались структурой самих генов. Но внутри ядра гены не существуют в «голом» виде, легко доступные для РНК-полимеразы или какого-либо другого белка, связывающегося с энхансером или промотором. Напротив, эукариотические хромосомы содержат столько же белка (по массе), сколько нуклеиновой кислоты, и этот ДНК-белковый комплекс называют хроматином. Доминирующими белками хроматина являются гистоны. Эти пять белков – HI, Н2А, Н2В, Н3 и Н4 представляют собой сильноосновные полипептиды с высоким содержанием положительно заряженных аминокислот лизина и аргинина. Другой гистон, Η5, обнаружен в клетках немногих типов (а именно в эритроцитах птиц и амфибий), в которых ДНК упакована предельно плотно. В этих клетках Н5 заменяет HI. Пять перечисленных главных гистонов имеются у всех представителей царств животных, растений и простейших, и в ходе эволюции изменились очень мало. Обычно такой консерватизм означает, что белки должны играть исключительно важную роль во всех изученных клетках. Ключевая функция гистонов заключается в упаковке ДНК в специфические спиральные структуры, называемые нуклеосомами Нуклеосома это основная единица структуры хроматина; она состоит из гистонового октамера (H2A, Н2В, Н3, H4)2. вокруг которого намотано два витка ДНК, составляющих примерно 140 пар оснований (рис. 12.22). Между нуклеосомами лежат другие 60 (или около того) пар оснований ДНК; эти «линкерные» нуклеотиды могут быть покрыты гистоном ΗI. Такая структура предполагается на основе опытов по расщеплению хроматина, когда он подвергается гидролизу небольшими количествами дезоксирибонуклеазы (ДНКазы) из определенных бактерий (Micrococcus). Эта обработка приводит к расщеплению ДНК по некоторым сайтам, и полученные фрагменты хроматина могут быть отделены друг от друга с помощью центрифугирования в градиенте плотности сахарозы (рис. 12.23). Когда ДНК экстрагируют из обработанного хроматина и разделяют в гелях, получается четкая картина. Вместо кусков случайного размера ДНК предстает расщепленной на фрагменты, кратные по длине 200
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 160_______________ ГЛАВА 12__________________________________________________________
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ______________________________ 161
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 162_______________ ГЛАВА 12________________________________________________________________________ парам оснований (Hewish, Burgoyne, 1973; Noll, 1974). Если эти фрагменты хроматина, обработанного ДНКазой, рассматривать в электронный микроскоп, то фрагмент, содержащий 200 пар оснований, виден как сферическое тело (глобула) с небольшим хвостом. Последовательность из 400 пар оснований представляется в виде двух связанных глобул, а последовательность из 600 пар оснований содержит три сферические тела, связанные друг с другом (Finch et al., 1975). На основе этих наблюдений был сделан вывод, что главная субъединица хроматина содержит около 200 пар оснований ДНК и что микрококковая ДНКаза расщепляет ДНК преимущественно между нуклеосомами. Роль гистонов в образовании этой структуры была продемонстрирована в опытах, в которых удалось реконструировать субъединицы хроматина в пробирке (Kornberg, Thomas, 1974). Смешивание гистонов H2А, H2В, H3 и H4 приводит к образованию только тетрамеров из H2А и H2В, а также тетрамеров из H3 и H4. Однако если к этой смеси гистонов добавить ДНК, то нуклеиновые кислоты и гистоны агрегируют с образованием нуклеосомной структуры с теми же самыми физическими и химическими свойствами, что и обычный хроматин. Таким образом, хроматин можно представить себе как нитку нуклеосомных бус, связанных между собой 10–100 парами оснований ДНК. Гистоновый октамер состоит из двух молекул каждого из гистонов H2А, H2В, H3 и H4 и приблизительно 140 пар оснований ДНК, которая намотана на белковую глобулу. Гистон H1 не представлен в нуклеосомах, но в зависимости от физиологического состояния клетки может быть связан с линкерной ДНК между нуклеосомами. Эти нуклеосомы в свою очередь упаковываются в плотные структуры, а H1, по-видимому, участвует в наматывании нуклеосом друг на друга, особенно при подготовке к клеточному делению (рис. 12.24) (Weintraub, 1984). Кроме того, по крайней мере в одном случае, показано, что определяемая H1 конформация нуклеосом ингибирует транскрипцию специфических генов в соматических клетках (Schlissel, Brown, 1984). 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|