
|
|
Дополнительные сведения и гипотезы: Молекулярная основа быстрой транскрипции рибосомных генов
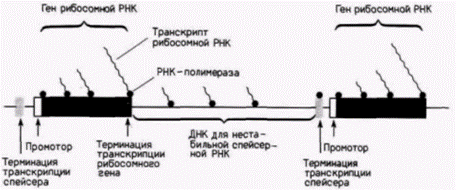
Для осуществления транскрипции необходимо взаимодействие РНК-полимеразы с ДНК. Однако далеко не каждый фрагмент ДНК подходит для этого (в противном случае транскрипция могла бы начаться где угодно и клетка заполнилась бы дефектными молекулами мРНК). Участок ДНК, специализированный для связывания РНК-полимеразы, называют промотором. В клетках эукариот содержатся РНК-полимеразы трех различных типов, каждый из которых характеризуется определенными свойствами и функцией (Rutter et al., 1976). РНК-полимераза I обнаруживается в области ядрышка в ядре и отвечает за транскрипцию больших рРНК, РНК-полимераза II транскрибирует предшественники мРНК, а РНК-полимераза III транскрибирует малые РНК, такие, как транспортные РНК, рибосомную 5S-PHK и другие небольшие последовательности РНК. Эти полимеразы имеют различную чувствительность к ионным условиям и антибиотикам. Их свойства представлены в табл. 11.1. Высокая эффективность транскрипции рибосомных генов может быть обусловлена структурой области спейсера между соседними генами рибосомной РНК. Как у большинства генов, промотор, присоединяющий РНК-полимеразу, лежит непосредственно перед (5’) кодирующей областью гена. Однако в отличие от большинства других генов у генов рРНК прямо перед промотором имеется последовательность ДНК, отвечающая за терминацию транскрипции (рис. 11.13). Эта структура (до сих пор обнаруженная только в рибосомных генах млекопитающих и амфибий) может концентрировать молекулы РНК-полимеразы на этих генах. У большинства генов при терминации транскрипции происходит высвобождение РНК и РНК-полимеразы в нуклеоплазму. РНК-полимераза может затем использоваться повторно, если найдет на ДНК новый промоторный сайт. Этот процесс, на который заведомо должно тратиться время, возможно, не осуществляется при транскрипции рибосомных генов. Вместо этого после отделения 40S-предшественника рРНК от гена РНК-полимераза остается на спейсере, синтезируя, вероятно, короткие нестабильные транскрипты (Labhart, Reeder, 1986). Как отмечалось ранее, непосредственно перед промотором находится специальный участок терминации транскрипции. Этот участок взаимодействует с промотором для обеспечения переноса РНК-полимеразы к соседнему гену (McStay, Reeder, 1986; Henderson, Sollner-Webb, 1986; Grummt et al., 1986). Поэтому, коль скоро РНК-полимераза начала транскрипцию рибосомного гена, она переносится затем от одного
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 120_______________ ГЛАВА 11___________ ____________
гена к другому без вхождения в пул несвязанных полимераз. Не исключено также, что свободно плавающие молекулы РНК-полимеразы I способны узнавать определенные участки ДНК в спейсерах и вовлекаться в транскрипцию рибосомной РНК. В этом случае спейсер будет действовать как «зона загрузки», накапливая новые полимеразы для транскрипции генов рибосомной РНК (Moss, 1983; Henderson, Sollner-Webb, 1986).
Известно, что при синтезе белков хориона у Drosophila melanogasler гены хориона также амплифицируются. Белки хориона синтезируются фолликулярными клетками яичника. Перед началом экспрессии генов хориона весь геном фолликулярных клеток проходит дополнительные циклы синтеза ДНК, так что количество ДНК достигает 16 гаплоидных наборов. После этой репликации гены белков хориона реплицируются избирательно еще около десяти раз. Эта амплификация происходит только в фолликулярных клетках яичника (Spradling, Mahowald, 1980).
Амплификация обусловлена дополнительными циклами репликации ДНК лишь в определенных областях генома (Spradling, 1981). Эти области ДНК характеризуются наличием нескольких ветвей, каждая из которых содержит амплифицированные гены хориона (рис. 11.14 и 11.15). В данном случае амплификация происходит, но гены остаются присоединенными к хромосомам. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|



 Гены хориона дрозофилы
Гены хориона дрозофилы