|
|
Клонирование у амфибий: исключения из ограничения потенцийМожно, однако, и по-иному объяснить ограничение потенций ядер в дифференцированных клетках. Перенося ядро дифференцированной клетки в цитоплазму зрелого яйца, мы тем самым заставляем его вернуться к теперь уже не свойственному ему физиологическому состоянию. У лягушек в период дробления ядра клеток делятся с очень высокой скоростью, тогда как в некоторых дифференцированных клетках они делятся редко или не делятся вообще. Неспособность к репликации ДНК быстро приводит к поломкам хромосом: такие хромосомные аномалии наблюдаются во многих клетках головастиков, развившихся из яиц, содержавших клонированные ядра. Джон Гёрдон и его коллеги, используя несколько измененную методику пересадки ядер, получили результаты, показывающие, что многие из ядер дифференцированных клеток остаются тотипотентными. Главное различие между опытами Гёрдона и опытами Бриггса и Кинга заключалось в выборе объекта исследований. Гёрдон изолировал ядра из клеток Xenopus laevis, южноафриканской шпорцевой лягушки. Эта лягушка (рис. 9.13) гораздо более
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. ТОЖДЕСТВО ГЕНОМОВ И ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ 75
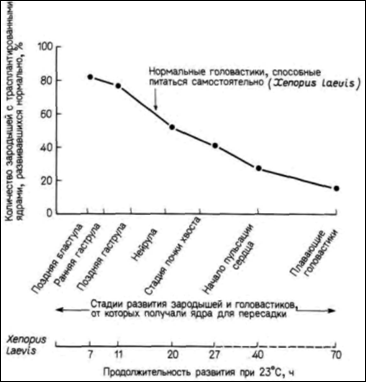
примитивна, чем Rana pipiens; y нее отсутствуют веки, среднее ухо и даже язык, столь характерный для позднее эволюционировавших видов Rana. Шпорцевая лягушка отличается также и по биологии развития. В отличие от леопардовой лягушки взрослая особь Xenopus способна регенерировать утраченные конечности; раннее развитие у нее протекает в три раза быстрее, чем у леопардовой лягушки. В частности, для достижения стадии хвостовой почки зародышу Rana pipiens требуется 80 ч, тогда как Xenopus laevis достигает той же стадии развития всего за 26 ч. Следовательно, ядра энтодермальных клеток на стадии хвостовой почки у шпорцевой лягушки имеют такой же возраст, как ядра из клеток ранней гаструлы у Rana (McKinnell, 1978). Гёрдон также обнаружил постепенную утрату потенций ядер по мере развития (рис. 9.14). Однако из этого правила выявились интересные исключения. Гёрдон переносил ядра из энтодермальных эпителиальных клеток кишечника, взятые у головастика Xenopus на стадии перехода к активному питанию, в активированные энуклеированные яйца. Ядра клеток донора имели генетический маркер одно ядрышко на клетку (линия 1-nu) вместо обычных двух (линия 2-nu). Из 726 пересаженных ядер только 10 оказались способными обеспечить развитие зиготы до стадии донора. Число ядер, обеспечивающих развитие, удалось повысить до 7% (Gurdon. 1962) с помощью метода серийных пересадок (переноса ядра из клеток кишечника в яйцо и после достижения яйцом-реципиентом стадии бластулы, последующего переноса ядер клеток бластулы в большее число яиц). В некоторых случаях ядра были способны реализовать информацию, достаточную для достижения стадии головастика и образования клеток всех линий нейронов, клеток крови и др. Кроме того, семь головастиков (развившихся после пересадки в яйца ядер, полученных клонированием двух исходных ядер) метаморфизировали и превратились в половозрелых взрослых лягушек (Gurdon, Uehlinger. 1966). Эти ядра были тотипотентными (рис. 9.15).
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 76________________ ГЛАВА 9_______________________________________________________________________________ Однако Кинг и его коллеги с сомнением отнеслись к опытам Гёрдона, считая, во-первых, что исследователи не приняли достаточных мер предосторожности, исключающих возможность использования вместо ядер кишечника ядер первичных половых клеток, которые, мигрируя, часто задерживаются в кишке, и. во-вторых, что эпителиальные клетки кишечника таких ранних, перешедших на активное питание головастиков могут не обладать свойствами вполне дифференцированных клеток. У них эти клетки еще содержат желточные пластинки (DiBerardino, King, 1967; McKinnell, 1978; Briggs, 1979). В ответ на эти критические замечания Гёрдон и его коллеги провели следующие опыты. Они культивировали эпителиальные клетки из кожных межпальцевых перепонок ноги взрослой особи Xenopus. Эти клетки были, безусловно, дифференцированными, потому что каждая из них содержала кератин – белок, характерный для клеток кожи взрослых особей. Когда ядра этих клеток пересаживали в энуклеированные яйца, ни одно из ядер не обеспечивало развития зародышей дальше нейрулы. Однако после клонирования ядер и их пересадки в энуклеированные яйца из таких яиц развивались многочисленные головастики: правда, все они погибали, не переходя на активное питание (Gurdon el al.. 1975). Сходную остановку развития наблюдали другие исследователи (Wabl et al., 1975), пытаясь получить взрослых лягушек при пересадке ядер лимфоцитов. Ди Берардино (DiBerardino, 1987) пришла к заключению, что «на сегодня еще не доказано, что ядра клеток хотя бы одного какого-либо специализированного типа или клеток взрослого организма являются тотипотентными». Результаты опытов по клонированию ядер у амфибий позволяют сделать два вывода. Во-первых, в них можно увидеть общее правило ограничения потенций в процессе развития. Это ограничение генетически детерминировано и характерно для ядер определенного типа клеток-доноров. Во-вторых, можно легко убедиться в том, что геном дифференцированной клетки обладает изумительной способностью инициировать образование всех типов клеток головастика. Ядро эритроцита лягушки, в котором никогда не происходит репликации ДНК (и синтеза мРНК), может претерпеть при серийных пересадках в общей сложности свыше 100 делений и все же сохранить способность обеспечивать развитие энуклеированного и активированного яйца в свободно плавающего головастика (Orr et al., 1986). Другими словами, даже если еще и ведутся споры о тотипотентности таких ядер, то в том, что они в высокой степени полипотентны, сомневаться не приходится. Ясно одно многие неиспользованные гены клеток кожи или крови могут реактивироваться и обеспечить образование нервов, желудка и сердца свободно плавающего головастика.
Как мы уже убедились ранее, обсуждая работы Ру и Дриша, детали экспериментальной методики могут сильно повлиять на получаемые результаты. Незначительное изменение процедуры клонирования ядер леопардовой лягушки привело к тому, что ядра, взятые даже на поздней стадии развития, обеспечивали развитие нормальной личинки. Салли Хеннен (Hennen, 1970) показала, что эффективность действия ядер донора можно увеличить, обработав их перед трансплантацией спермином или охладив яйца после пересадки, чтобы дать ядру время для адаптации к цитоплазме яйца. Обрабатывая таким способом ядра энтодермальных клеток, взятых у зародыша леопардовой лягушки на стадии хвостовой почки, и пересаживая затем эти ядра в энуклеированные яйца, Хеннен в 60% случаев получала нормальных личинок (в контрольных опытах процент таких личинок был равен нулю). Однако обработка ядер, взятых из дифференцированных клеток кожи взрослых лягушек, спермином не приводила к развитию взрослых особей из энуклеированных яиц, в которые пересаживали эти ядра. Клонирование человека из дифференцированных клеток, похоже, стало задачей некоторых журналистов и писателей (писатели задавались целью воспроизводить важных политических деятелей, таких, как Гитлер или Кеннеди, а журналисты возмечтали распространить эту методику на атлетов и кинозвезд). Из предыдущего обсуждения очевидно, что клонирование полностью развитого индивидуума из дифференцированных клеток невероятно трудное дело, да и результаты этих опытов не вполне убедительны. Даже у амфибий ядра дифференцированных клеток, взятые у взрослого животного, не способны после их пересадки в активированные и энуклеированные яйца обеспечить развитие из этих яиц взрослого животного. Но даже если бы и удалось вырастить взрослых лягушек из яиц, в которые были пересажены ядра дифференцированных клеток, то этот результат
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ТОЖДЕСТВО ГЕНОМОВ И ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ________________ 77
нельзя экстраполировать на человека. Не говоря уже об этических проблемах, при работе с человеческим материалом исследователь сталкивается с многочисленными трудностями. Если у самки амфибий одновременно созревают сотни яиц, то у женщины ежемесячно образуется очень мало зрелых яиц. Кроме того, цитоплазма из яйцеклетки женщины может существенно отличаться от цитоплазмы яйцеклетки лягушки по своей способности воспринимать сигналы, исходящие из ядра клетки, которая находится на более продвинутой стадии развития. Пересадки ядер были впервые осуществлены у мышей. С этой целью у одной зиготы удаляли оба пронуклеуса и замещали их пронуклеусами. взятыми из другой зиготы (McGrath, Solter, 1983). Процедура этого опыта показана на рис. 9.16. Сначала одноклеточных зародышей инкубируют в среде с цитохалазином и колхицином, чтобы ослабить микрофиламенты и микротрубочки цитоскелета. Затем зародыша удерживают на месте с помощью присасывающей пипетки, а пипеткой для энуклеации прокалывают прозрачную оболочку (zona pellucida) яйца. Цитоплазматическую мембрану клетки энуклеирующей пипеткой не прокалывают, а лишь нажимают ею на область, где находятся пронуклеусы, и втягивают внутрь пипетки этот участок мембраны вместе с прилежащей к нему цитоплазмой и пронуклеусами (А). Затем пипетку оттягивают и тем самым цитоплазму с пронуклеусами отделяют от яйца. Эта цитоплазма окружена плазматической мембраной (Б). Пипетку с пронуклеусами, окруженными мембраной, погружают в каплю, содержащую инактивированный вирус Сендай, который вызывает слияние мембран. После всасывания некоторого количества вируса пипетку приближают к другой зиготе, из которой таким же образом были удалены пронуклеусы. Прокалывают прозрачную оболочку и вводят пронуклеусы вместе с окружающей их мембраной в перивителлиновое пространство между оболочкой яйца и его плазматической мембраной (В). Затем зародыш инкубируют при температуре 37°С до момента слияния мембраны яйца-хозяина и донора ядер (Г). Таким образом два пронуклеуса донора попадают в цитоплазму хозяина. Через пять дней культивирования зародыши достигают стадии бластоцисты, и их можно имплантировать в матку приемной псевдобеременной взрослой самки. Родившийся мышонок имеет фенотип мыши, служившей донором ядер. Свыше 90% энуклеированных зигот мыши, получивших ядра из других зигот, успешно развивались до стадии бластоцисты. Когда же в энуклеированные зиготы пересаживали ядра 4-клеточных
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 78________________ ГЛАВА 9___________________________________________________________________________
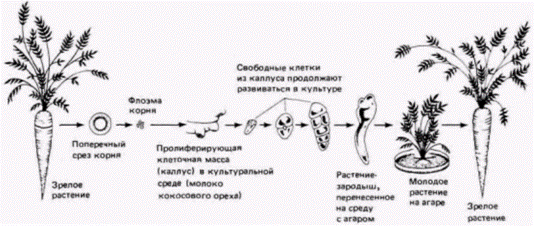
зародышей, ни одна зигота (из 81) не достигала стадии бластоцисты. Точно так же ядра из клеток 8-клеточного зародыша и из клеток внутренней клеточной массы были не способны поддерживать развитие до предимплантационных стадий (McGrath. Solter. 1984). В отличие от морских ежей и амфибий у мышей ядра ранних бластомеров (которые, как известно, являются тотипотентными) не способны обеспечить полное развитие. Эти опыты не удаются, по-видимому, из-за того, что ядра бластомеров не могут правильно функционировать в цитоплазме зиготы. Таким образом, вряд ли стоит серьезно относиться к идее (высказанной в сентябре 1984 г. в журнале «National Examiner») о клонировании второго Элвиса Пресли из его дифференцированной клетки. Однако свойства ранних бластомеров не у всех млекопитающих одинаковы: разные виды сильно различаются по времени активации и имплантации зародышей в матку. Недавно появилось сообщение (Willadsen, 1986) о том, что, используя модифицированную методику трансплантации ядер, автор получил ягнят из зигот, в которые были пересажены ядра бластомеров 8-клеточного зародыша. У мыши ядра бластомеров не способны направлять развитие зиготы, а у овцы они, по-видимому, могут это делать. Если указанное сообщение подтвердится, его результаты будут иметь огромное значение для сельского хозяйства. Случаинесомненной способности ядер клеток взрослого организма обеспечивать развитие другого взрослого организма обнаружены только у растений. Эта способность чрезвычайно четко продемонстрирована на клетках моркови и табака. В 1958 г. Стюард и его коллега разработали методику, позволившую получить из дифференцированной ткани корня моркови целое новое растение (рис. 9.17). Из корня моркови изолировали маленькие кусочки флоэмы и вращали в больших колбах, содержащих молоко кокосового ореха (которое на самом деле является его жидким эндоспермом). Оно содержит различные факторы и питательные вещества, необходимые для роста растений, а также гормоны, необходимые для их дифференцировки. В этих условиях клетки флоэмы делятся и формируют неорганизованную ткань, называемую каллусом. Непрерывное вращение вызывает выталкивание отдельных клеток из каллуса в суспензию. Из этих единичных суспендированных клеток образуются корнеподобные узелки, которые продолжают расти все время, пока остаются в суспензии. Если эти узелки перенести на среду, уплотненную агаром, то из них развиваются остальные части растения: в конце концов образуются целые растения моркови, способные к размножению (Steward et al., 1964; Steward, 1970). Весь процесс развития от единичной клетки до цветущего растения в условиях вращающейся культуры наблюдать невозможно, однако Вазил и Хильдебрандт (Vasil, Hildebrandt, 1965) проследили эти события, изолировав единичные клетки табака и наблюдая за их развитием или непосредственно, или с помощью цейтраферной киносъемки. Как и из клеток моркови, из клеток табака образовывались растеньица, которые могли расти и давать семена. Клетки табака дают нам еще один пример тотипотентности ядер. Все гены, необходимые для образования целого растения, имеются в ядре дифференцированной клетки. Однако растения и животные развиваются по-разному. Вегетативное размножение отводками (т.е. частями растения, которые. получая питательные вещества, регенерируют недостающие части) представляет собой обычный прием в практике сельского хозяйства. В отличие от животных (у которых половые клетки обособляются
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ТОЖДЕСТВО ГЕНОМОВ И ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ___________________ 79
очень рано в развитии в виде отчетливой клеточной линии) у растений в норме гаметы образуются из соматических клеток. Поэтому нет ничего удивительною в том, что единичная клетка может давать начало другому типу клеток и сформировать генетически однородный клон (от греч. klon, что означает «ветвь»). Итак, мы можем сделать вывод, что дифференциальная утрата генов не является причиной дифференцировки. Ядра дифференцированных клеток содержат большую часть, а возможно, и все гены зиготы: эти гены экспрессируются в соответствующих условиях. Следовательно, процесс дифференцировки включает в себя селективную экспрессию разных частей генома. Нам предстоит более глубокое исследование проблемы постоянства генома и дифференциальной экспрессии генов с привлечением методов молекулярной биологии и клонирования генов. В следующей главе обсуждение этой проблемы будет продолжено на уровне современной молекулярной биологии. ЛИТЕРАТУРА
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|


 Дополнительные сведения и гипотезы: Клонирование
Дополнительные сведения и гипотезы: Клонирование