|
|
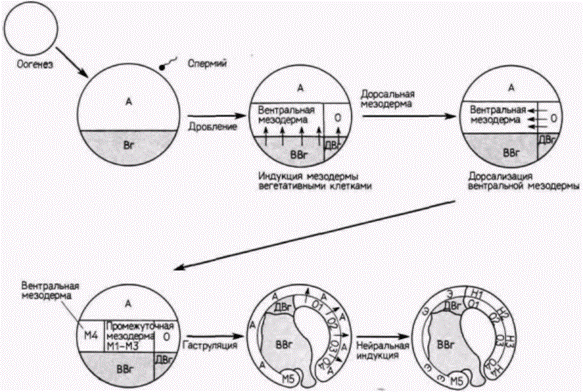
Влияние индукции на активность геновСогласно приведенной выше модели, эктодерма и энтодерма специфицируются как таковые автономно. Лишь мезодерма приобретает свой статус путем индукции. Эта модель появилась на свет
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 58 ГЛАВА 8
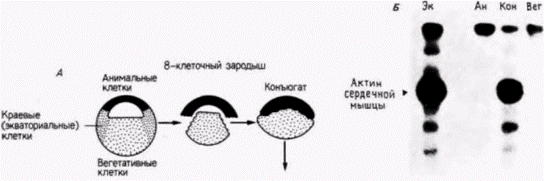
благодаря недавним исследованиям, в которых удалось идентифицировать клеточные РНК (такие методики будут изложены подробнее в гл. 10). Сарджент и др. (Sargent et al., 1986) диссоциировали ранние бластулы шпорцевой лягушки (от 32- до 128-клеточной стадии) на составляющие их клетки, удаляя оболочку оплодотворения и помещая зародышей в культуральную среду, не содержащую ионов Са и Mg. Они решили выяснить, будут ли эти диспергированные клетки по прошествии соответствующего времени синтезировать мРНК, специфичные для данного зародышевого листка. Результаты исследований показали, что гены, специфичные для энтодермы или синтеза специфического белка кишечника ) и для эктодермы (для синтеза определенного белка цитоскелета), активируются и с них транскрибируются новые мРНК даже в диспергированных клетках. Это открытие позволило предположить, что клетки энтодермы регулируются автономно содержащимися в них цитоплазматическими факторами. Однако гены, специфические для мезодермы (гены α-актина), не активируются в изолированных клетках. Когда зародышей диссоциировали даже на 128-клеточной стадии, в потомках диссоциированных клеток не удалось обнаружить мРНК α-актина. Но этот ген можно было активировать, если клеткам давали возможность формировать скопления и таким образом взаимодействовать между собой. Отсюда следует, что экспрессия генов мезодермы требует, по-видимому, взаимодействия по меньшей мере двух типов клеток. Наблюдения Ньюкопа были также подтверждены Гёрдоном и др. (Gurdon et al., 1985). Если морфологические исследования Ньюкопа правильны, то вегетативные клетки, помещенные непосредственно под клетки анимального полушария (которые в норме дают эктодермальные ткани), должны индуцировать в них активность α-актинового гена. У зародыша на стадии средней бластулы Гёрдон и его коллеги отрезали клетки краевой зоны и рекомбинировали клетки анимального полюса с массой вегетативных клеток (рис. 8 22). Они обнаружили, что в клетках анимального полюса началась транскрипция специфичной для мезодермы мРНК α-актина. Вегетативные клетки индуцировали включение специфического для мезодермы гена в презумптивной эктодермальной ткани. Одновременно эти взаимодействия вызывали утрату специфической активности эктодермальных генов (Sargent et al., 1986).
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 60________________ ГЛАВА 8___________________________________________________________________ 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|