
|
|
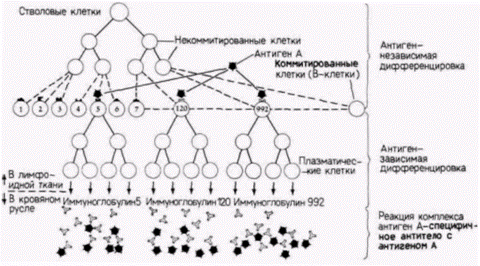
Нарушение стабильности геномов: изменения в генах лимфоцитовИспользование методов молекулярной биологии позволило выявить интересное исключение из сформулированного выше правила; исключение это – дифференцировка лимфоцитов. В соответствии с общим правилом неиспользуемые гены присутствуют в дифференцированных клетках и сохраняют способность к функционированию. Набор генов одинаков во всех тканях. Геном же каждого типа лимфоцитов отличается от генома любых других типов клеток в организме, в том числе и от геномов других типов лимфоцитов. Примером такого рода являются В-лимфоциты клетки, которые синтезируют антитела. Антитела образуются, когда чужеродный субстрат (антиген) приходит в контакт с В-лимфоцитами, находящимися в лимфатических узлах и селезенке. Еще до контакта с антигеном в каждом из
1 В последнее время широко используется слияние клеток или их фрагментов в электрическом поле. – Прим. ред.
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 94________________ ГЛАВА 10______________________________________________________________________________
покоящихся В-лимфоцитов синтезируются молекулы антител, однако из клеток они не выделяются, а встраиваются в клеточные мембраны лимфоцитов. Каждый B-лимфоцит продуцирует антитела, которые узнают один и только один антиген. Таким образом, антитела, узнающие белковую оболочку вирусов полиомиелита, не будут узнавать холерный токсин, мембрану клетки Е. coli или вирус гриппа. После того как связанные с мембраной антитела присоединяют молекулы определенного антигена, В-лимфоцит многократно делится и дифференцируется в плазматическую клетку, секретирующую антитела (рис. 10.17 и 10.18). (Механизм этой дифференцировки будет подробно рассмотрен в гл. 16.) К размножению и секреции антител стимулируются только те В-лимфоциты, которые обладают способ-
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. _____________ ТОЖДЕСТВО ГЕНОМОВ И ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ_______________________ 95
ностью связывать данный антиген. В соответствии с этой моделью, названной теорией клональной секреции (Burnett, 1959), каждый B-лимфоцит приобретает присущую ему специфичность до контакта с антигеном. Иными словами, из миллионов различных видов антител, которые каждый В-лимфоцит может производить, он «выбирает» только один вид и размещает антитела этой специфичности на своей клеточной поверхности. Механизм этой селекции по специфичности антител включает образование новых генов в ходе дифференцировки B-лимфоцитов. Белок антитела на клеточной поверхности состоит из двух пар полипептидных субъединиц (рис. 10.19): двух идентичных тяжелых цепей и двух идентичных легких цепей. Цепи связаны друг с другом дисульфидными связями. Специфичность молекулы иммуноглобулина (т.е. избирательность связывания с вирусом полиомиелита, клеткой Е. coli или какими-либо другими молекулами) определяется последовательностью аминокислот в вариабельной области. Эта область формируется из амино-концов одной тяжелой цепи и одной легкой цепи. Вариабельные области молекул иммуноглобулинов присоединены к константным областям, которые придают антителу его эффекторные свойства. Например, константная область тяжелых цепей молекул поверхностных иммуноглобулинов удерживает эти белки в клеточной мембране. ФОРМИРОВАНИЕ ГЕНОВ ЛЕГКИХ ЦЕПЕЙ АНТИТЕЛ. Гены для легких и тяжелых цепей лимфоцитов разделены на сегменты. Гены легких цепей состоят из трех сегментов (рис. 10.20). Первый сегмент гена кодирует вариабельную (V) область легкой цепи. Она содержит около 300 различных последовательностей, которые кодируют в общей сложности пер-
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 96________________ ГЛАВА 10______________________________________________________________________________
вые 97 аминокислот легкой цепи антитела. Второй сегмент, J-область, состоит из четырех или пяти возможных последовательностей ДНК для последних 15—17 остатков вариабельной части легкой цепи антитела. Третий сегмент определяет константную (С) область легкой цепи. В ходе развития В-лимфоцита одна из трехсот V-областей и одна из пяти J-областей комбинируются друг с другом и образуют вариабельную часть гена антитела. Это достигается перемещением последовательности V-области к последовательности J-области перестройки, которая сопровождается удалением промежуточной ДНК. О такой перестройке гена впервые сообщалось в работе Хозуми и Тонегавы (Hozumi, Tonegawa, 1976). Эти исследователи выделили ДНК из зародышей мыши и опухолевых B-клеток, секрети-
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. ______________ ТОЖДЕСТВО ГЕНОМОВ И ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ___________________ 97 рующих легкие цепи 1. Каждый из двух препаратов ДНК обрабатывали рестриктазой BamHI,которая расщепляет в ДНК последовательность ГГАТЦЦ, где бы она ни находилась. В результате были получены серии фрагментов ДНК. Размеры каждого фрагмента определялись длиной участка ДНК между двумя сайтами расщепления. Полученные фрагменты ДНК наносили на край желеобразного плоского геля, через который пропускали электрический ток (рис. 10.21). При миграции ДНК к положительному электроду меньшие по размеру фрагменты двигались быстрее, чем ее крупные фрагменты, за счет чего достигалось эффективное разделение фрагментов по размерам. Гель с распределенной по нему ДНК разрезали на кусочки, каждый из которых содержал фрагменты ДНК определенного размера2. Из каждого кусочка элюировали ДНК и денатурировали ее. Часть этой ДНК гибридизовали с радиоактивной РНК, которая кодирует полную легкую цепь и была изолирована из исходных опухолевых B-клеток. Другую часть гибридизовали с радиоактивной РНК, которая кодирует лишь С-область легкой цепи (фрагмент мРНК с 3'-конца). ДНК из клеток зародышей связывалась с мРНК легкой цепи в двух зонах геля. ДНК в первой зоне имела молекулярную массу около 6 млн. дальтон, тогда как ДНК во второй зоне – 3,9 млн. дальтон. При гибридизации ДНК зародышей мыши с мРНК для С-области легкой цепи эта РНК связывалась лишь с ДНК из зоны с молекулярной массой 6 млн. Таким образом, в зародышах мыши С-область кодируется внутри фрагмента ДНК с молекулярной массой 6 млн. (между сайтами BamHI). тогда как V-область кодируется внутри фрагмента с массой 3,9 млн. (рис. 10.22). Принципиально иной результат был получен для ДНК из опухолевых лимфоцитов. Единственная зона ДНК, связывающая мРНК легкой цепи, имела молекулярную массу 2,4 млн. Кроме того, эта зона связывала фрагмент мРНК, кодирующий С-область легкой цепи. Оказалось, что обе области, С и V, кодируются одним и тем же фрагментом ДНК! Простейшее объяснение, которое было многократно подтверждено в других лабораториях и с помощью иных методов (см. Brack et al., 1978; Bernard el al., 1978; Seidman ei al., 1979), заключается в том, что два фрагмента гена, один из которых кодирует С-область легкой цепи и другой – специфичную V-область легкой цепи, соединились вместе, образовав новый ген. Новый ген возник в ходе развития лимфоцита. Предложенная авторами схема образования такого гена показана на рис. 10.23. ФОРМИРОВАНИЕ ГЕНОВ ТЯЖЕЛЫХ ЦЕПЕЙ АНТИТЕЛ. Гены тяжелых цепей антител содержат даже боль-
1 Опухолевые клетки были использованы потому, что они синтезируют огромное количество определенного иммуноглобулина (и мРНК для этого иммуноглобулина). 2Размер ДНК в каждом кусочке геля определяли по миграции фрагментов ДНК с известной длиной на параллельной дорожке.
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 98________________ ГЛАВА 10______________________________________________________________________________
шее число сегментов, чем гены легких цепей. Сегменты гена тяжелой цепи включают V-область (200 различных последовательностей для первых 97 аминокислот). D-область (10-15 различных последовательностей для 3-14 аминокислот) и J-область (четыре последовательности для последних 15–17 аминокислот V-области). Следующий фрагмент – это С-область. Вариабельная область тяжелой цепи формируется в результате присоединения одной V-последовательности и одной D-последовательности к одной из J-последовательностей (рис. 10.24. А, Б). Эта VDJ-последовательность вариабельной час-
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ТОЖДЕСТВО ГЕНОМОВ И ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ________________ 99 ти далее присоединяется к первой С-области тяжелойцепи, специфичной для антител, которые могут быть встроены в плазматическую мембрану. Таким образом, молекулы антител формируются двумя генами, возникающими в ходе антигеннезависимой стадии развития В-лимфоцита. Эти молекулы встраиваются в клеточную мембрану и служат рецепторами антигенов. ПЕРЕКЛЮЧЕНИЕ КЛАССОВ В ГЕНАХ ТЯЖЕЛЫХ ЦЕПЕЙ. После стимуляции антигеном В-клетка делится и дифференцируется в плазматическую клетку секретирующую антитела. Вначале антитела, синтезируемые этими клетками, содержат ту же С-область, что и ранее. Однако при дальнейшем синтезе антител С-область может измениться. Исходную С-область называют константной мю-областью (Cμ). Следующая С-область та, что содержится в секретирующих антителах, может остаться μ-областью (хотяи с модификацией, которая обеспечивает секрецию), но может также стать гамма(γ)эпсилон(ε)- или альфа(α)-константной областью. Таким образом, один и тот же вариабельный участок тяжелой цепи может соединяться вначале с константной μ-областью, а позже, например, с константной γ-областью. Это явление называется переключением классов. (Класс тяжелых цепей антител определяет характер их функционирования: μ- и γ-цепи стимулируют лизис, агглютинацию или разрушение антигена макрофагами: ε-цепи вызывают воспалительный процесс, а α-цепи обеспечивают выделение антител в слизь, слезы, слюну и молоко.) Переключение классов осуществляется транслокацией полного сегмента вариабельной области гена (VDJ) из положения перед константной μ-областью в участок, примыкающий к константной γ-, ε- или α-области (рис. 10.24. В, Г). Этот процесс заключается в делеции сегмента константной μ-области гена из генома (Davis et al., 1980; Cory et al., 1980; Rabbits et al., 1980; Yaoita, Honjo, 1980). Таким образом, геном плазматической клетки существенно отличается от генома любой другой клетки. Во-первых, в нем была создана последовательность вариабельной области гена посредством объединения различных сегментов ДНК. В клетках всех других органов эти сегменты ДНК разделены, а в В-лимфоцитах и плазматических клетках они собраны вместе. Во-вторых, во многих плазматических клетках часть генома (а именно ДНК из константной μ-области тяжелых цепей) элиминируется из ядер. Определенная часть генома утрачивается в ходе развития плазматической клетки: создаются новые гены, тогда как другие разрушаются. Какие выводы можно сделать? Имеющиеся данные свидетельствуют о том, что ядра дифференцированных клеток действительно сохраняют основную часть своей генетической информации в форме, которая допускает ее экспрессию в соответствующих условиях. Тем не менее очевидно, что по крайней мере при дифференцировке клеток одного типа происходит некоторая утрата генетического материала. Пока у нас нет возможности выяснить, каково разнообразие необратимых генетических изменений, которые могут происходить в процессе развития животных Однако известные нам сведения о перестройке генов в лимфоцитах указывают на то, что необратимые генетические потери являются следствием клеточной дифференцировки, а не ее причиной.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|






 (см. след. страницу)
Рис. 10.22. Протокол и результатыопыта Хозуми и Тонегавы. ДНК из эмбриональных клеток мыши и В-клеток по отдельности гидролизовали рестриктазой Ват HI, фракционировали с помощью электрофореза и элюировали из гелей. Каждый элюированный препарат ДНК после денатурации гибридизовали или с полной мРНК. кодирующей вариабельную и константную области легкой цепи иммуноглобулина, или с фрагментом мРНК, кодирующим только константную область легкой цепи белка (3'-конца). В случае эмбриональной ДНК вариабельная и константная области легкой цепи белка были обнаружены в двух различных фрагментах ДНК (константная область располагалась на фрагменте с молекулярной массой 3,9 х 106. а вариабельная область – на фрагменте ДНК с массой 6 х 106). В опухолевых лимфоцитах константная и вариабельная области располагались вместе на одном фрагменте ДНК, имеющем молекулярную массу 2,4 х 106. (По Hozumi, Tonegawa, 1976.)
(см. след. страницу)
Рис. 10.22. Протокол и результатыопыта Хозуми и Тонегавы. ДНК из эмбриональных клеток мыши и В-клеток по отдельности гидролизовали рестриктазой Ват HI, фракционировали с помощью электрофореза и элюировали из гелей. Каждый элюированный препарат ДНК после денатурации гибридизовали или с полной мРНК. кодирующей вариабельную и константную области легкой цепи иммуноглобулина, или с фрагментом мРНК, кодирующим только константную область легкой цепи белка (3'-конца). В случае эмбриональной ДНК вариабельная и константная области легкой цепи белка были обнаружены в двух различных фрагментах ДНК (константная область располагалась на фрагменте с молекулярной массой 3,9 х 106. а вариабельная область – на фрагменте ДНК с массой 6 х 106). В опухолевых лимфоцитах константная и вариабельная области располагались вместе на одном фрагменте ДНК, имеющем молекулярную массу 2,4 х 106. (По Hozumi, Tonegawa, 1976.)


