|
|
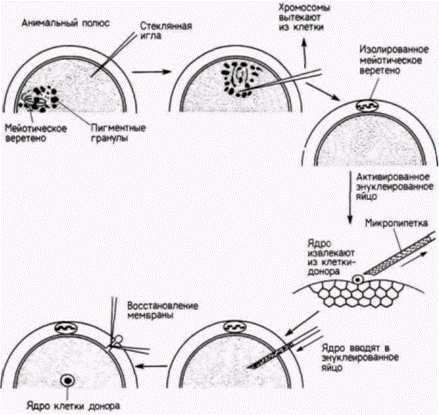
Клонирование у амфибий: ограничение потенций клетокОкончательно решить вопрос о том, является ли сужение функций ядра дифференцированной клетки необратимым, было бы можно, проверив способность этого ядра индуцировать образование дифференцированных клеток любого другого типа. В 1938 г. Ганс Шпеман предложил, по его словам, «фантастический» опыт, чтобы ответить на вопрос о том, действительно ли геномы в разных клетках идентичны. Для этого следует имплантировать ядро какой-либо дифференцированной клетки в яйцо, собственное ядро которого предварительно было удалено. Если любое пересаженное ядро идентично ядру зиготы, то оно должно обеспечить полное развитие организма. Однако для проведения такого опыта необходимо было прежде всего разработать три методики: I ) методику энуклеации яиц-реципиентов без их разрушения. 2) методику изоляции неповрежденных ядер; 3) методику переноса таких ядер в яйцо без повреждения и ядра, и яйца. Указанные методики были разработаны Робертом Бриггсом и Томасом Кингом. Эти исследователи комбинировали энуклеацию яйца с его партеногенетической активацией. Если яйцеклетку леопардовой лягушки (Rana pipiens) уколоть стеклянной иглой, то в ней начнут происходить все цитологические и биохимические изменения, связанные с оплодотворением: разрушаются кортикальные гранулы, перемещается внутренняя цитоплазма, а вблизи анимального полюса завершается мейоз.
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 72________________ ГЛАВА 9_____________________________________
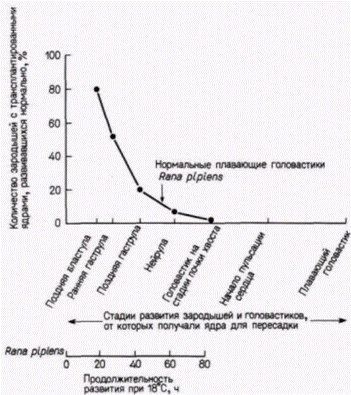
Положение мейотического веретена в яйце легко определить, поскольку на анимальном полюсе пигментные гранулы удаляются от него и прокол яйца в этом месте приводит к вытеканию мейотического веретена вместе с хромосомами наружу (рис. 9.9). Теперь яйцо является и активированным (так как процессы, необходимые для начала развития, свершены), и одновременно энуклеированным. В энуклеированные яйца переносят ядра из клеток-доноров. Для этого клетку разрывают, с помощью микропипетки захватывают ее ядро и вводят его в яйцо-реципиент. Вместе с ядром в яйцо попадает и некоторое количество окружающей его цитоплазмы клетки-донора, но соотношение цитоплазмы донора и реципиента столь ничтожно (1:105), что вряд ли она оказывает какое-либо влияние на исход опыта. В 1952 г. Бриггс и Кинг (Briggs, King, I952) показали, что ядра, взятые у зародыша на стадии бластулы, будучи перенесенными в цитоплазму яйцеклетки, обеспечивают развитие полностью сформированного головастика. Еще раньше Шпеман продемонстрировал, что клетки бластулы не детерминированы и, следовательно, их ядра тотипотентны. Поэтому если система переноса ядер работает, то ядра бластулы должны быть способны обеспечить полное развитие. Именно это и наблюдается в опытах по пересадке ядер бластулы. Шестьдесят процентов всех имплантированных ядер оказались способными направлять развитие яиц до стадии свободно плавающего головастика; все эти головастики были диплоидными (результат, подтверждающий, что их ядра происходили из ядра клетки-донора). Следовательно, система переноса ядер работает и ее можно использовать для изучения потенций ядра (рис. 9.10). Что произойдет, если в активированное энуклеированное яйцо пересадить ядро из клетки зародыша, находящегося на более продвинутой стадии развития? Результаты опытов Кинга и Бриггса (King, Briggs, 1956) суммированы на рис. 9.11. Из этого рисунка видно, что большинство ядер из клеток
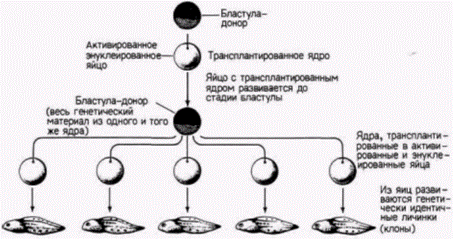
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ТОЖДЕСТВО ГЕНОМОВ И ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ___________________ 73 бластулы способны обеспечить развитие яиц-реципиентов до стадии нормальных свободно плавающих головастиков, однако в ядрах, взятых от доноров, находящихся на более поздних стадиях развития, эта способность резко ограничивается. Ни одно из ядер соматических клеток,взятых на стадии хвостовой почки, не могло дать информацию, необходимую для развития нормального зародыша. Однако, когда на той же стадии брали ядра половых клеток (в норме дающих после оплодотворения начало целому организму), в 40% случаев были получены бластулы, способные к дальнейшему развитию (Smith, 1956). Следовательно, соматические клетки, становясь детерминированными и дифференцированными, по-видимому, утрачивают способность обеспечивать полное развитие организма. Было показано, что ограничение потенций ядер по мере развития является стабильным и тканеспецифическим признаком. Данные в пользу этого получали в следующих опытах. Число энтодермальных ядер, взятых на стадии поздней гаструлы, увеличивали путем серийных пересадок. С этой целью одно ядро переносили в энуклеированное яйцо; развившийся в результате такой процедуры зародыш имел на стадии бластулы тысячи идентичных ядер. Ядра этой бластулы снова переносили в энуклеированные яйца и таким образом получали много копий первоначального ядра, что позволяло количественно оценить его потенции. Такаяметодика называется клонированием ядер(рис. 9.12). При трансплантации неклонированных ядер наблюдается большая изменчивость в способности ядер, полученных от разных доноров, обеспечивать развитие. Некоторые ядра обеспечивают весь путь развития до стадии свободно плавающего головастика, тогда как другие - только до стадии аномальной гаструлы. Кинг и Бриггс считали эту изменчивость в разных клонах нормальной. Однако они обнаружили, чтов пределах одного клона все ядра обладают одинаковыми потенциями. Каждый клон имел «характерный» фенотип, и часто стадия, на которой останавливалось развитие особей, полученных в результате пересадки ядер потомков одного клонированного ядра энтодермальной клетки, была сходной. Это наблюдалось и при трансплантации ядер, клонированных в течение нескольких поколений. Кроме того, если личинки развивались с какими-либо дефектами, то эти дефекты у всех личинок были одинаковыми. Все личинки с клонированными ядрами имели энтодермальные структуры (а именно, кишку), но у них отсутствовали некоторые производные мезодермы или эктодермы. По-видимому, ядра из клеток энтодермы пригодны для формирования энтодермы, но их способность к формированию эктодермы или мезодермы ограничена. Сходная утрата потенций была обнаружена и в ядрах клеток эктодермы (DiBerardino, King, 1967). Аномальные головастики имели превосходно дифференцированные нейральные структуры, но у них отсутствовали энтодермальные производные. Таким образом, прогрессивное ограничение потенций ядер по мере развития является, по-видимому, общим правилом. Не исключено, что ядра разных дифференцированных клеток отличаются друг от друга.
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 74________________ ГЛАВА 9______________________________________________________________________________ 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|