|
|
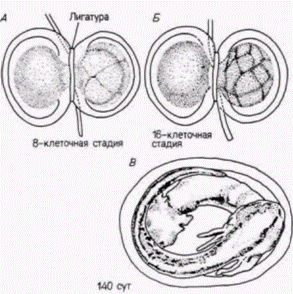
Ганс Шпеман: прогрессивная детерминация эмбриональных клетокВ предыдущем разделе главы были приведены данные о регуляционном типе развития. Мы отметили два главных аспекта регуляции: 1) потенция изолированных бластомеров в эмбриогенезе шире, чем их нормальная судьба, и 2) бластомеры. перемещенные в другую область зародыша, развиваются согласно их новому положению. Оба этих явления характерны для ранних стадий дробления морского ежа. Однако впоследствии бластомеры морского ежа становятся коммитированными к их различным проспективным значениям. Гёрстадиусу удалось связать ограничение потенций с ориентацией плоскости делений дробления, поскольку бластомеры могли регулировать развитие лишь до тех пор, пока они имели достаточно материала как из анимальной, так и из вегетативной частей яйца. В 1918 г. Ганс Шпеман (H. Spemann) из Фрайбургского университета обнаружил, что сходная ситуация наблюдается и в яйце тритона. Опыты, с помощью которых он и его коллеги анализировали это явление в течение более чем 20 лет. заложили основу большей части наших знаний о физиологии развития и обусловили присуждение Шпеману Нобелевской премии в 1935 г. Шпеман, подобно Ру и Дришу, решил проверить гипотезу Вейсмана и с помощью остроумной методики доказал, что ядра ранних бластомеров тритона идентичны, т. е. каждое из них способно обеспечить развитие целой личинки. Пользуясь волоском ребенка в качестве лигатуры, он перевязывал им яйцо тритона вскоре после оплодотворения в плоскости первого деления дробления. Затем он несколько стягивал петлю так, что все деления ядер происходили лишь в одной из половин. Наконец на стадии 16 бластомеров одно ядро смогло проскользнуть через перетяжку в безъядерную половину. Дробление начиналось и в этой половине, а петлю, накинутую на яйцо. Шпеман стягивал все сильнее, пока не разделял яйцо на две изолированные половины. В результате развивались два зародыша-близнеца, причем один был немного старше другого (рис. 8.11). Результаты этого опыта позволили Шпеману сделать вывод, что ядра ранних зародышей амфибий идентичны и каждое способно обеспечить развитие целого организма. В этом отношении бластомеры амфибий были сходны с бластомерами морских ежей. Однако, когда Шпеман проделал сходный опыт с перетягиванием яйца также лонгитудинально, но перпендикулярно к плоскости первого деления дробления (т.е. разделял яйцо не на левую и правую половины, а на будущую спинную и брюшную стороны), он получил совершенно другой результат! По обе стороны от лигатуры ядра продолжали делиться, но лишь из клеток одной стороны образовалась нормальная личинка. Из другой половины возникала только неорганизованная масса ткани, названная Шпеманом Bauchstück «кусок живота». Эта масса тканей представляла собой шарик эпидермальных клеток (эктодерма), содержащий внутри кровь и мезенхиму (мезодерма) и клетки кишки (энтодерма), но в нем не было дорсальных структур, таких, как нервная система, хорда или сомиты (рис. 8.12). Почему описанные выше два опыта дали разные результаты? Не могло ли это быть вызвано тем, что при первом делении яйца, когда плоскость деления проходила перпендикулярно к нормальной плоскости первого деления дробления, некоторые цитоплазматические вещества неравномерно распределялись по двум половинам? К счастью, яйцо тритона оказалось очень удобным для получения ответа на этот вопрос. Как уже говорилось в гл. 2 и 4, в яйцах амфибий после оплодотворения происходит резкое смещение кортикального слоя цитоплазмы и у некоторых видов амфибий такое движение приводит к образованию серого серпа в области, прямо противоположной месту проникновения спермия в яйцо. Кроме того, плоскость первого деления дробления обычно делит эту область поровну между двумя бластомерами. из которых, если их отделить друг от друга, развиваются две нормальные личинки. Однако, если плоскость первого деления отклоняется от середины серого серпа (в редких случаях спонтанно или в опыте, в котором исследователь перетягивает яйцо волосяной петлей перпендикулярно плоскости нормального деления), то материал
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 50________________ ГЛАВА 8_______________________________________________________________________________
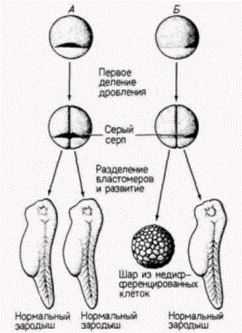
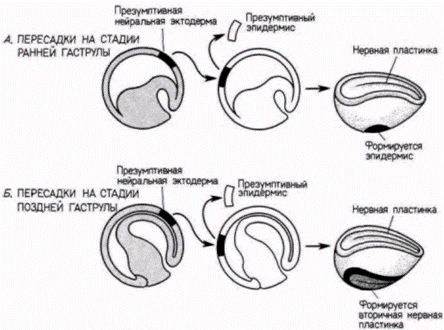
серого серпа может попасть только в один из двух бластомеров. Шпеман обнаружил, что когда эти два бластомера разделены полностью, нормально развивается только тот бластомер, который содержит материал серого серпа. Отсюда следует, что в области серого серпа содержится, по-видимому, какой-то фактор, весьма существенный для правильного развития зародыша. Но как этот фактор действует? Какую роль играет в нормальном развитии? Наиболее важный ключ к решению этого вопроса дает судьба области серого серпа. Показано, что из нее образуются клетки, которые инициируют гаструляцию и формируют спинную губу бластопора. Клетки спинной губы бластопора (см. гл. 4) запрограммированы, чтобы инвагинировать внутрь зародыша и, таким образом, начинать гаструляцию и формирование архентерона. Поскольку вес будущее развитие амфибий зависит от взаимодействия клеток, перемещающихся в процессе гаструляции. Шпеман предположил, что материал серого серпа играет решающую роль в инициации гаструляции и что в период гаструляции в развитии зародыша происходят кардинальные изменения. В 1918 г. Шпеман показал, что в период гаструляции действительно резко меняется потенция клеток. Он обнаружил, что клетки ранней гаструлы еще не детерминированы к их конечной дифференцировке, тогда как на стадии поздней гаструлы судьба клеток уже определена. Шпеман произвел пересадки тканей на стадии ранней гаструлы у двух различно пигментированных видов тритонов (рис. 8.13). Если участок презумптивного эпидермиса трансплантировали в область будущей нервной пластинки, то он превращался в нервную ткань. Λ если клетки презумптивной нервной пластинки пересаживали в область будущего эпидермиса живота, то они становились эпидермальными (табл. 8.2). Следовательно, клетки ранней гаструлы тритона не были коммитированы к специфическому типу дифференцировки. Их проспективные потенции были все еще шире их проспективного значения. Принято говорить о зависимом развитии этих клеток, поскольку их судьба зависит от их локализации в зародыше. Однако, когда такие же самые гетеропластические (межвидовые) пересадки были осуществлены на стадии поздней
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. ______________________ ПРОГРЕССИВНАЯ ДЕТЕРМИНАЦИЯ______________________________________________________________ 51
гаструлы. Шпеман получил совершенно другой результат. Вместо того, чтобы регулировать собственную дифференцировку в соответствии со своим новым положением, трансплантированные клетки обнаружили независимое (или автономное) развитие. Их проспективное значение уже было зафиксировано, и клетки развивались независимо от своего нового положения в зародыше. Презумптивные нервные клетки теперь образовывали нервную ткань даже в том случае, когда их помещали в область будущего эпидермиса, а презумптивный эпидермис формировал кожу, даже будучи помещенным в область презумптивной нервной трубки. Следовательно, за время, прошедшее от стадии ранней гаструлы до стадии поздней гаструлы. потенции этих групп клеток сузились, так что они
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 52________________ ГЛАВА 8________________________________________________________________________ смогли следовать только одному своему специфическом) пути дифференцировки. Эти клетки теперь можно назвать детерминированными. Под детерминацией подразумевается коммитирование (предназначение) клеток к тому, чтобы в конечном счете дифференцироваться именно в этот, а не в какой-либо иной специфический тип. Поэтому на стадии поздней гаструлы клетки анимального полюса на той стороне, где закладывается спинная губа бластопора, коммитированы (детерминированы) к образованию нервной ткани в любом месте, куда бы они ни попадали (включая чашку Петри). Их развитие уже не поддается регуляции, и они не могут превратиться в клетки других типов. Следует отметить, что критерием для определения состояния детерминации служит поведение клеток в условиях опыта. Никаких явных изменений в клетках не происходит, и еще не видно никаких признаков их дифференцировки. Молекулярные основы детерминации остаются одной из главных и пока нерешенных проблем биологии развития. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|