|
|
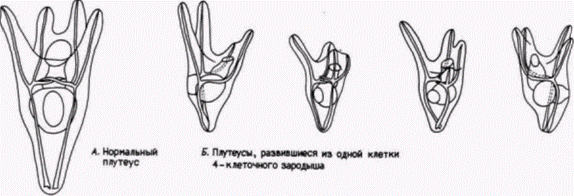
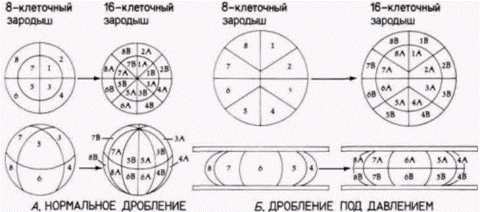
Ганс Дриш: регуляционное развитиеНикто не приветствовал этот экспериментальный подход к эмбриологии больше, чем другой ученик Геккеля – Ганс Дриш (H. Driesch). Дриш ставил перед собой цель объяснить развитие с позиций законов физики и математики. Первые его исследования были сходны с исследованиями Ру. Методически опыты Ру были опытами с нанесением дефектов; они отвечали на вопрос, как будут развиваться оставшиеся бластомеры, когда часть их разрушена. Дриш (1892) задумал расширить эти исследования, проделав опыты с изоляцией бластомеров. Он отделял друг от друга бластомеры морского ежа энергичным встряхиванием (или позже помещением их в бескальциевую морскую воду). К удивлению Дриша, каждый из бластомеров двуклеточного зародыша развивался в полную личинку. Точно также, если Дриш разделял бластомеры 4и 8-клеточных зародышей, то некоторые из клеток образовывали целые личинки, называемые плутеусами (рис. 8.3). Этот результат поразительно отличался от того, что предсказывали Вейсман или Ру. Вместо самодифференцировки в будущую часть зародыша каждый бластомер мог регулировать свое развитие и давать начало целому организму. Это явление было названо регуляционным развитием. Регуляционное развитие было продемонстрировано Дришем и в других опытах. У морских ежей плоскости двух первых делений дробления являются меридиональными, тогда как третье деление проходит по экватору яйца, разделяя зародыш на четыре верхние и четыре нижние клетки (см. рис. 3.3). Дриш (1893) изменял направление третьего деления, слегка сжимая ранний зародыш между двумя стеклянными пластинками; в результате третье деление также проходило в меридиональной плоскости, как и первые два. Если затем давление ослабляли, то четвертое деление дробления было экваториальным. Эта процедура вызывала перегруппировку ядер, так что ядро, которое в норме должно было оказаться в области, предназначенной формировать энтодерму, теперь попадало в область презумптивной эктодермы. Ядра, которые должны были обра-
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 42________________ ГЛАВА 8_______________________________________________________________________________
зовывать дорсальные структуры, обнаруживались в вентральных клетках (рис. 8.4). Если бы в данном случае происходила сегрегация ядерных детерминантов, то у развившегося из этих клеток зародыша все структуры должны были располагаться в причудливом беспорядке. Однако Дриш получил из этих зародышей нормальных личинок. Результаты этих опытов оказались очень важными как для эмбриологии, так и лично для Дриша Во-первых. Дриш показал, что «проспективная потенция» изолированного бластомера (т.е. тот тип клеток, который мог из него произойти) шире, чем его «проспективная судьба» (т.е. те типы клеток, которые должны формироваться из этого бластомера при неизменном ходе его развития). Согласно же Вейсману и Ру. проспективная потенция и проспективная судьба бластомера должны быть идентичными. Во-вторых. Дриш пришел к выводу, что зародыш морского ежа представляет собой «гармоничную эквипотенциальную систему». Система эта гармонична потому, что все ее потенциально независимые части функционируют вместе, формируя единый организм. В-третьих, судьба ядра зависела исключительно от его положения в зародыше. Дриш (1894) выдвинул гипотезу о серии событий, продвигающих развитие вперед посредством взаимодействия ядра и цитоплазмы: «Поскольку каждая клетка содержит ядро, она на протяжении онтогенеза несет в себе всю сумму зачатков; поскольку она содержит специфическое
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ПРОГРЕССИВНАЯ ДЕТЕРМИНАЦИЯ__________________________________________________ 43
цитоплазматическое клеточное тело, это позволяет ей реагировать только на специфические воздействия... Когда в какой-либо клетке активируется ядерный материал, цитоплазма этой клетки, которая влияла на ядро, сама в свою очередь оказывается под влиянием ядра и изменяется; таким образом, устанавливается основа для нового элементарного процесса, который является не только результатом, но также и причиной изменений». Эта удивительно современная концепция ядерноплазменного взаимодействия и равноценности ядер оказалась не по силам Дришу. Рассматривать зародыш как физическую машину было уже невозможно, ведь зародыш можно было разделить на части, каждая из которых была способна воссоздать целый организм. Другими словами. Дриш пришел к убеждению, что развитие не может быть объяснено физическими силами. И чтобы объяснить, каким образом происходит развитие, он был вынужден призвать на помощь жизненную силу – энтелехию («внутреннюю силу, направляющую к цели»). В сущности, зародыш должен быть наполненным внутренним духом и мудростью, чтобы осуществить свои цели, несмотря на препятствия, которые ставят на его пути эмбриологи. Неспособный объяснить свои результаты с позиций физики его времени. Дриш отказался от изучения физиологии развития, стал профессором философии и продолжал провозглашать витализм вплоть до своей смерти в 1941 г. Различия в опытах Ру и Дриша суммированы в табл. 8.1. Различия между опытами по нанесению дефектов и изоляции, а также важность взаимодействий, обеспечиваемых разрушенными бластомерами. были выяснены в опытах Мак-Клендона (McClendon. 1910), показавших, что изолированные бластомеры лягушки ведут себя точно так же. как изолированные бластомеры морского ежа. Следовательно, мозаичное развитие бластомеров лягушки в исследованиях Ру было артефактом, полученным в опытах с нанесением дефекта. Что-то в мертвом бластомере или на нем все еще информировало живые клетки о его существовании. Мы видели также, что ранние бластомеры млекопитающих обладают регуляционным типом развития. Как уже обсуждалось в гл. 3. каждый изолированный бластомер внутренней клеточной массы у мыши способен генерировать целую фертильную мышь. Такие факты, как слияние двух и более ранних зародышей мыши в один нормальный зародыш (см. рис. 3.3) или рождение идентичных близнецов, также свидетельствуют о том. что ранние бластомеры млекопитающих способны к регуляции. Поэтому, хотя Вейсман и Ру были пионерами исследований в области физиологии развития, их предположение о том, что причиной дифференцировки является сегрегация ядерных детерминантов, вскоре было опровергнуто. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|