|
|
Цитоплазматическая спецификация у зародышей оболочниковЭЛЕКТРОННОЕ ОГЛАВЛЕНИЕ
БИОЛОГИЯ РАЗВИТИЯ
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.
С. Гилберт БИОЛОГИЯ РАЗВИТИЯ
«Мир» Москва
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. ББК 28.0 Г47 УДК 57 Гилберт С. Г47 Биология развития: В 3-х т. Т. 2: Пер. с англ.-М.: Мир. 1994.-235с. ил. ISBN 5-03-001832-8 Фундаментальное учебное и справочное пособие по относительно новой, быстро развивающейся дисциплине биологии развития. На русском языке выхолит в 3-х томах. Второй том посвящен вопросам клеточной дифференцировки. Для эмбриологов, молекулярных биологов, генетиков, цитологов и других специалистов-биологов, а также студентов биологических факультетов. ББК 28.0 Редакция литературы по биологии Издание выпущено в счет дотации, выделенной комитетом РФ по печати
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.
Я считаю вероятным, что в зародышевых клетках существуют тонкие внутренние различия, которые предопределяют их последующую трансформацию в детерминированные структуры, не различия, которые являются простыми потенциями, присутствующими в зародышевых клетках, а действительные материальные различия, столь тонкие, что мы пока еще не в состоянии выявить их. ВИРХОВ (1858) Изучая период дробления, мы приближаемся к источнику, дающему начало прогрессивно разветвляющимся потокам дифференцировок. которые завершаются в конце концов почти тихими заводями индивидуальными клетками взрослого организма. ДЖАСТ (1939)
Введение Каждый многоклеточный организм представляет собой сложный набор специализированных клеточных типов. Например, эритроциты и лейкоциты отличаются не только друг от друга, но и от клеток сердечной мышцы, которая обеспечивает циркуляцию форменных элементов крови по всему телу. Они отличаются также от имеющих отростки нейронов, проводящих нервные импульсы от мозга к сердцу, и от железистых клеток, выделяющих в кровь гормоны. В табл. 7.1 представлен далеко не полный список специализированных клеточных типов с указанием характерных продуктов их жизнедеятельности и основных функций. Процесс развития специализированных клеточных типов из одного оплодотворенного яйца называется дифференцировкой. Этому явному изменению в биохимии и функции клеток предшествует процесс, называемый детерминацией, в течение которого определяется судьба клеток. Детерминация может осуществляться двумя разными способами. Первый заключается в цитоплазматической сегрегации детерминирующих молекул в период дробления, в результате чего качественно различные области цитоплазмы зиготы попадают в разные дочерние клетки. Второй способ детерминации - эмбриональная индукция – заключается во взаимодействии клеток или тканей, определяющем судьбу одного или обоих участников этого взаимодействия. Как будет показано ниже, в развитии любого организма в разной степени участвуют оба механизма. В этой главе основное внимание будет уделено опытам, свидетельствующим о существовании цитоплазматической сегрегации: следующая глава будет посвящена детерминации путем индукции (прогрессивной детерминации). Преформация и эпигенез Любая трактовка возникновения разных типов дифференцированных клеток из оплодотворенного яйца должна объяснить I) постоянство морфологии каждого типа (т.е. почему из куриного яйца всегда вылупляется цыпленок, но не крокодил) и 2) разнообразие частей тела каждого организма. И в самом деле, одной из основных характеристик развития является то. что каждый вид воспроизводит свои специфические черты эмбриогенеза. Развитие включает в себя экспрессию наследственных свойств вида. В XVII столетии понятия развитие и наследст-
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 6_________________ ГЛАВА 7______________________________________________________________
венность были объединены в гипотезе, которая получила название гипотеза преформации. В соответствии с этой гипотезой считалось, что все органы взрослого организма в миниатюре представлены в спермин или (гораздо чаще) в яйце. Следовательно, организмы не «развиваются», а, скорее, «развертываются». Эта гипотеза основывалась как на научных данных, так и на философских концепциях (Gould, 1977; Roe. I981). Во-первых, считалось, что. поскольку все органы предобразованы, зародышевое развитие - это всего лишь рост существующих структур, а не формирование новых. Из этого вытекало, что эмбриональное развитие не нуждалось в действии какой-либо внешней таинственной силы. Во-вторых, точно так же, как взрослый организм был преформирован в первичных половых клетках, другое поколение было преформировано внутри первичных половых клеток первого преформированного поколения. Эта гипотеза, названная теорией вложения (инкапсуляции), давала уверенность в том, что вид всегда будет оставаться постоянным. Некоторые микроскописты утверждали, что видят полностью сформированных миниатюрных человечков в спермии или в яйце, однако главные защитники этой гипотезы Альбрехт фон Галлер (Haller) и Шарль Боннэ (Bonnet) знали, что системы органов развиваются с разной скоростью и что эмбриональные структуры не обязательно находятся в том же месте, в каком эти структуры находятся у новорожденного. В распоряжении преформистов еще не было клеточной теории, чтобы они могли предусмотреть нижний предел размеров их преформированных организмов, и они не смотрели на пребывание человечества на Земле как на нечто потенциально бесконечное. Боннэ (Bonnet, 1764) говорил: «Природа может создать любую малость», а человеческий род существует лишь ограниченное время, которое отпущено ему между Сотворением мира и днем Страшного суда. Это утверждение не противоречило науке того времени, подтверждая принцип французского математика и философа Рене
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ДЕТЕРМИНАЦИЯ ПОСРЕДСТВОМ ЦИТОПЛАЗМАТИЧЕСКОЙ СПЕЦИФИКАЦИИ__________________ 7 Декарта о бесконечной делимости механической природы, делимости, начатой Богом, в которую он далее не вмешивался. Теория преформации была консервативна, поскольку из нее вытекало, что каких-либо изменений в последовательных поколениях происходить не может. Принципиальный недостаток этой теории заключался в том. что с ее помощью невозможно было объяснить изменчивость, которая к тому времени была известна из очень небольшого числа генетических данных. Было известно, например, что от белого и черного родителей могут рождаться дети с промежуточным цветом кожи, что невозможно, если наследственность и развитие обусловливаются природой либо спермия. либо яйца. В более строго контролируемых опытах немецкий ботаник Кёльрейтер (Kölreuter. 1766) получил гибриды табака, имеющие признаки обоих видов. Более того, скрещивая гибрид либо с мужским, либо с женским родителем, он смог после нескольких поколений «возвратить» гибрид обратно к одному или к другому родительскому типу. Таким образом, стало ясно, что признаки потомства представляют собой некую смесь родительских признаков. Помимо этого, теория преформизма не могла объяснить случаи рождения уродов и детей с явными отклонениями (например, с шестью пальцами на конечностях) у нормальных родителей. Нужна была другая гипотеза, и она была предложена - это гипотеза эпигенеза. В соответствии с ней постулировалось, что каждый взрослый организм развивается заново из недифференцированного состояния. Этот взгляд на развитие, имевший философские корни, идущие еще от воззрений Аристотеля, был возрожден немецким эмбриологом, работавшим в Санкт-Петербурге. Каспаром Фридрихом Вольфом (К. F. Wolff). Тщательно наблюдая за развитием куриного зародыша, Вольф показал, что части зародыша развиваются из тканей, не имеющих аналогов во взрослом организме. Так. например, можно было проследить, что сердце и кровеносные сосуды (которые, согласно гипотезе преформизма, должны присутствовать с самого начала, чтобы обеспечивать рост зародыша) у каждого зародыша развиваются заново. Сходным образом можно было видеть, что пищеварительная трубка возникает в результате сворачивания первоначально плоской ткани. Это последнее наблюдение было детально описано Вольфом, заявившим (1767): «Если тщательно взвесить все данные об образовании кишечника таким способом, то, я полагаю, практически не остается сомнений в правильности теории эпигенеза». Однако чтобы объяснить создание организма заново в каждом поколении, Вольф был вынужден постулировать некую неизвестную силу, vis essentialis («жизненную энергию»), которая, действуя подобно силе тяжести или магнетизму, должна была организовывать эмбриональное развитие. Следовательно, если с позиций преформизма можно было легче объяснить непрерывность поколений, то с позиций эпигенеза легче объяснить изменчивость и результаты прямых наблюдений над формированием органов. Немецкий философ Иммануил Кант (1724-1804) и его коллега биолог Иоганн Фридрих Блуменбах (1752-1840) попытались примирить эти противоположные гипотезы. Стремясь построить научную теорию происхождения рас. Блуменбах постулировал существование механической целенаправленной силы, названной им Bildungstrieb («формообразующее стремление»). Существование этой силы, утверждал он. может быть доказано не только теоретически, но и экспериментально. Например, гидра, будучи разрезанной, регенерирует ампутированные части посредством перераспределения существующих элементов. Следовательно, в данном случае наблюдается действие некоей целесообразной организующей силы и эта сила заключена в самом организме. Блуменбах полагал, что Bildungstrieb наследуется через половые клетки. Таким образом, развитие могло происходить эпигенетически благодаря предетерминированной силе, наследуемой с веществом зародыша (Cassirer, 1950; Lenoir. 1980). Кроме того, эта сила могла изменяться, и в качестве примера такого изменения приводили вариант левозакрученной спирали раковины улитки. От этой гипотезы, согласно которой преформированные инструкции направляют эпигенетическое развитие, не так далеко до взглядов некоторых современных биологов, считающих, что «полное описание организма уже записано в яйце» (Brenner, 1979). Однако вплоть до начала двадцатого столетия, когда вновь были открыты работы Менделя, последовательной генетической теории, в которой нашли бы место такие идеи о наследственной изменчивости, не существовало, и каждый ученый мог свободно строить свои предположения о механизмах наследования признаков в развитии. Французские тератологи Попытки найти гипотезу, объясняющую одновременно постоянство вида и эпигенетический характер развития, привели к созданию современной эмбриологии. Поиски такой гипотезы были предприняты в двух различных направлениях. Одно из этих направлений, разрабатываемое во Франции, основывалось на анатомических аспектах тератогенеза. Ученые пытались выявить ошибки в эмбриогенезе, которые приводили к рождению детей с аномалиями развития. Другое направление поисков
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 8_________________ ГЛАВА 7______________________________________________________________________________ такой гипотезы было сосредоточено в Германии, и здесь внимание было сфокусировано на физиологии развития. Обе исследовательские группы начали экспериментировать на зародышах с целью изучить реакции развивающегося организма на различные вмешательства (Churchill. 1973: Fischer, Smith, 1984). Тератологические опыты французов начались в 20-х годах прошлого века с исследований Этьена Жоффруа Сент-Илера и его сына Исидора. Эти исследователи пытались доказать, что врожденные аномалии являются результатом нарушенного развития плода, а не преформированных отклонений. Поэтому они надеялись искусственно вызвать аномалии развития, изменяя условия инкубации куриных яиц. В большинстве случаев эти попытки были безуспешными (из-за грубой методики зародыши либо продолжали развиваться нормально, либо погибали), но они подготовили почву для проведения более тонкого анализа в работах Дареста (Dareste, 1877). Дарест проделал тысячи опытов и проследил возникшие у кур аномалии, начиная с ранних стадий развития. Однако куриный зародыш оказался плохим объектом для изучения самых ранних стадий эмбриогенеза. Чтобы выяснить, будут ли нарушения на ранних стадиях развития влиять на структуры взрослого организма, следовало использовать другое животное. В 1886 г. Лоран Шабри (Chabry, 1887) начал изучать тератогенез на более доступном зародыше оболочников. Это был удачный выбор, потому что зародыши оболочников быстро превращаются в личинку с относительно небольшим числом клеточных типов. Шабри собирался вызывать специфические нарушения, отделяя скальпелем определенные бластомеры дробящегося зародыша оболочников. Он обнаружил, что каждый бластомер ответствен за образование специфического набора тканей личинки. Если удалить какие-либо бластомеры, то у личинки будут отсутствовать как раз те структуры, которые в норме из них формируются. Кроме того, он обнаружил, что если изолировать определенные группы клеток зародыша, то из них формируются характерные структуры без связи с другими клетками. Следовательно, каждая из клеток у оболочников развивается, по-видимому, автономно. Такой способ развития часто называют мозаичным, потому что зародыш представляет собой как бы мозаику самодифференцирующихся частей. Цитоплазматическая спецификация у зародышей оболочников Более поздние исследования показали, что зародыш оболочников почти точно соответствует представлению о нем как о «мозаике самодифференцирующихся частей», конструируемой на основе информации, которая была накоплена в цитоплазме ооцита. По мере того как зародыш делится, в различные клетки включаются разные участки цитоплазмы. Полагают, что эти разные цитоплазматические участки содержат морфогенетические детерминанты, контролирующие детерминацию (коммитирование) данной клетки к образованию клеток определенного типа. Изучение детерминации клеток у оболочников в значительной степени облегчалось тем обстоятельством, что у некоторых видов цитоплазма яйца сразу после его оплодотворения сегрегируется на ряд по-разному окрашенных областей. (Это можно видеть на цветной таблице на внутренней стороне обложки книги.) Конклин (Conklin, 1905) описал, как эти окрашенные области распределяются по разным бластомерам. Первое деление дробления разделяет яйцо на правую и левую половины, являющиеся зеркальным отражением одна другой. Каждое последующее клеточное деление на правой и левой стороне происходит синхронно. Проследив судьбу каждого бластомера асцидии Styela partita, Конклин пришел к поразительному заключению, что каждая окрашенная область имеет свою особую судьбу (рис. 7.1). Желтый серп цитоплазмы дает начало мышечным клеткам; из серого экваториального серпа образуются хорда и нервная трубка; прозрачная анимальная цитоплазма становится эпидермисом личинки; из содержащей желток серой вегетативной области яйца возникает кишка личинки. Ревербери и Минганти (Reverberi, Minganti, 1946) изучали детерминацию у оболочников в серии экспериментов по изоляции бластомеров; эти авторы также наблюдали самодифференцировку каждого изолированного бластомера и остальной части зародыша. Результаты одного из опытов показаны на рис. 7.2. Если 8-клеточный зародыш разделить на четыре части по два бластомера в каждой (правая и левая стороны эквивалентны), то мозаичная детерминация их является правилом. Задняя анимальная пара бластомеров дает начало эктодерме, задняя вегетативная пара продуцирует энтодерму, мезенхиму и мышечную ткань, как это и следовало ожидать, исходя из карты презумптивных зачатков. Однако развитие нейральных структур является исключением. Клетки, образующие нервную систему, происходят из двух передних квадрантов – анимального и вегетативного, но ни один из них по отдельности не дает начало нервным клеткам. Если эти передние пары вновь соединяют вместе, то возникают ткани мозга и пальпы (прикрепительные сосочки). Другими словами, даже в таком строго детерминированном зародыше, как зародыш оболочников, осуществляются некоторые индукционные взаимодействия между бластомерами. И действительно, Ортолани (Ortolanι. 1959) показал, что эта
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ДЕТЕРМИНАЦИЯ ПОСРЕДСТВОМ ЦИТОПЛАЗМАТИЧЕСКОЙ СПЕЦИФИКАЦИИ______________________ 9
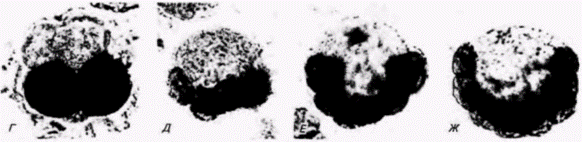
область эктодермы не детерминирована к состоянию «нейральности» вплоть до стадии 64 клеток, непосредственно предшествующей началу гаструляции. Таким образом, хотя большая часть тканей у этих зародышей детерминируется сразу же после оплодотворения, некоторые ткани претерпевают прогрессивную детерминацию. В 1973 г. Уиттейкер (Whittaker, 1973) получил убедительные биохимические данные, свидетельствующие о цитоплазматической сегрегации тканевых детерминантов у оболочников. Он окрашивал клетки на присутствие или отсутствие в них фермента ацетилхолинэстеразы. Этот фермент содержится только в мышечной ткани личинки и участвует в возникновении способности мышц реагировать на повторные нервные импульсы. Результаты исследований Конклина и других авторов, касающиеся судьбы потомства каждого из бластомеров, или клеточных линий (рис. 7.2 и 7.3), показали, что только одна пара бластомеров (задняя вегетативная. В4.1) у 8-клеточного зародыша способна дать начало мышечной ткани. Когда Уиттейкер удалял эти две клетки и культивировал их по отдельности, они образовывали мышечную ткань, которая положительно окрашивалась на ацетилхолинэстеразу. Из остальных 6/8 зародыша возникала личинка, лишенная мышц и не обнаруживавшая сколько-нибудь явной холинэстеразной активности, (рис. 7.4)1. Помимо этого Уиттейкер останавливал развитие зародышей оболочников на разных стадиях, обра-
1 Было показано (Deno et al., 1984; Nishida, 1987), что по меньшей мере у некоторых видов оболочников пары бластомеров b4.2 и A4.1 также дают начало мышечным клеткам. Однако это можно увидеть только на целых зародышах, а не на изолированных бластомерах. Эти мышечные клетки, происходящие не из пары бластомеров В4.1, по-видимому, образуются в результате индукции (Whittaker, 1987; Meedel et al, 1987).
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 10________________ ГЛАВА 7______________________________________________________________________________
батывая зародышей цитохалазином В. Это вещество связывается с микрофиламентами. предотвращая тем самым цитокинез (деление цитоплазмы клеток), и не влияет на нормальное деление ядер. В результате все дальнейшее развитие происходит в пределах популяции тех клеток, которые имелись к моменту добавления цитохалазина. После того как у зародышей было полностью блокировано дробление, им давали возможность развиваться какое-то время и затем окрашивали на присутствие ацетилхолинэстеразы. Результаты оказались поразительными. Их сравнение (рис. 7.5) с картой клеточных линий (рис. 7.3) показало полное совпадение данных. Способностью к образованию мышечных клеток первоначально обладали оба бластомера. Однако к 4-клеточной стадии способность синтезировать ацетилхолинэстеразу сохранялась только у вегетативных бластомеров. V 8-клеточного зародыша такие клетки формировались только из двух задних вегетативных бластомеров. Как известно, именно эти клетки формируют большую часть мышцы хвоста у личинки оболочника (Meedel et al.. 1987; Nishida, 1987). Клетки, потомки которых, как было показано, синтезировали ацетилхолинэстеразу, оказались клетками, предназначенными образовывать мышцы. Кроме того, синтез фермента в изолированных клетках происходил точно в то же время, в которое он появился в нормальных зародышах (Satoh. 1979). Когда в 8-клеточном зародыше асцидии цитоплазму из бластомера В4.1 (дающего начало мышцам) переносили в формирующий эктодерму бластомер
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 12________________ ГЛАВА 7______________________________________________________________________________
В4.2. из него помимо нормальных эктодермальных клеток возникали еще и мышечные (рис. 7.6; Whittaker, 1982). Напротив. Тунг и др. (Tung et al., 1977) показали, что если ядра личинки трансплантировать в энуклеированные фрагменты яиц. то вновь образующиеся клетки имеют структуры, типичные для клеток - доноров цитоплазмы, а не для клеток, являющихся донорами ядер. Отсюда мы можем заключить, что определенные детерминанты, содержащиеся в цитоплазме, вызывают образование определенных тканей. Эти морфогенетические детерминанты (или морфогены), по-видимому, действуют посредством избирательной активации (или инактивации) специфических генов. Следовательно, детерминация бластомеров и активация определенных генов контролируются пространственной локализацией морфогенетических детерминантов в цитоплазме яйца. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|


 Sinauer Associates, Inc. · Publishers
Sunderland, Massachusetts
Sinauer Associates, Inc. · Publishers
Sunderland, Massachusetts