|
|
Сердцевина, 6 — стерильная часть, 7 — спороносная часть,8 — спорангии, 9 — центральнаячасть нуцеллуса, 10 — интегумент, 11 — плюска(купула), 12 — пыльцевая камера Л КЛАСС САГОВНИКОВЫЕ (CYCADOPSIDA) Ныне живущая группа голосеменных растений. Геологическая история их началась еще до конца карбона (340 млн. лет назад), а временем расцвета были юрский и меловой периоды (195—61 млн. лет назад). Происходят они, вероятно, от семенных папоротников. Листья их широкие и перистые, реже дважды перистые. Устьица простогубые (гаплохейльные). Стебли имеют толстую сердцевину и относительно слаборазвитую и рыхлую вторичную древесину и толстую кору. Такой тип строения стебля называют маноксилъным (от греч. manos — рыхлый, неплотный и xylos — древесина). Спорофиллы собраны в однополые стробилы. Семязачатки с пыльцевой камерой. Сперматозоиды с многочисленными жгутиками. Архегонии развиты. В настоящее время представлен одним порядком — Cycadales, одним семейством — Cycadaceae, 9 родами и 100 видами в тропических и субтропических областях.
Род саговник (Cycas) насчитывает 16 видов. Распространен от Восточной Азии до Северо-Восточной Австралии и на о. Мадагаскар. Ныне живущие саговники являются остатками некогда широко распространенной группы растений. Ископаемые остатки саговников встречаются в отложениях триаса и юры. Тогда они были господствующими
Современные саговники— деревья с колонновид-ным стеблем высотой от 6 до 20 м или клубневидным, невысоким, скрытым в земле (рис. 41). У Zamia polygama, произрастающей на Кубе, стебель очень маленький — 3 см высотой и 2 см в диаметре.
Листья стеблевого происхождения, макрофиллия, простоперистые, как у пальм, реже — дважды—трижды перистые до 2 м длиной, жесткие, вечнозеленые, в молодом состоянии улиткообразно свернутые, как у папоротников. Стебель маноксильного типа, имеет сердцевину с крахмалом, за ней идет узкое кольцо древесины, затем следуют камбий, тонкий слой флоэмы и кора. В коре имеются слизевые ходы со слизью (сходство с мараттиевыми папоротниками). Корень саговников длинный, разветвленный. Для саговниковых характерным является наличие боковых поверхностных корней, растущих вверх. У поверхности земли или над землей они обильно ветвятся, образуя коралловидные или клубневидные скопления, которые располагаются у основания ствола. В парен- химе корней поселяются азотфиксирующие бактерии, сине-зеленые водоросли (анабена) и в межклетниках — гифы грибов. Боковые корни саговников вступают в множественный симбиоз с бактериями, водорослями и грибами. Спорофиллы собраны в шишки. Саговники — двудомные растения. На вершине стебля мужского экземпляра располагаются мужские шишки длиной 50—70 см. На оси шишки расположены микроспорофиллы (рис. 42). На нижней стороне микроспорофиллов расположены микросорусы из 2—4 микроспорангиев. Микроспорангий вскрывается продольной трещиной. В микроспорангиях развивается большое количество эллипсоидальных мелких спор. Микроспоры без воздушных мешков, прорастают. После первого деления микроспоры возникает одна маленькая клетка, соответствующая вегетативной части мужского гаметофита (по-гречески prothallium). Поэтому ее называют проталли-алъной клеткой. Более крупная клетка получила название антериди-алъной инициалы. Она делится еще раз и образует две специализированные клетки. Одна из них формирует антеридий и называется антеридиальной. Вторая клетка развивает гаусторию, поэтому ее называют гаусториалъной. В таком состоянии они попадают на семязачаток (мегаспорангий), где идет дальнейшее развитие мужского заростка. Мегаспорофиллы в небольшом числе расположены на верхушке стебля, чередуясь с вегетативными листьями. Перистые мегаспорофиллы сходны с вегетативными листьями, но меньше по размерам и окрашены в желтый или красный цвет. В нижней части мегаспорофилла, на его разветвлениях расположены шесть семязачатков по три с каждой стороны мегаспорофилла. Семязачатки крупные, до 5—6 см длины. Развитие семязачатка происходит следующим образом. На ме-гаспорофилле возникает нуцеллус в виде небольшого бугорка из одинаковых клеток. По мере его роста от основания нуцеллуса вырастает интегумент. В верхней части нуцеллуса образуется пыльцевая камера в виде углубления. Она заполнена слизью, выступающей за пределы семязачатка. Интегумент окружает нуцеллус. Он мощный, состоит из трех слоев: наружного и внутреннего мясистого и среднего каменистого. На верхушке интегумент вытянут в виде трубочки в микропиле (пыльцевход). В верхней части нуцеллуса выделяется одна крупная клетка — материнская клетка мегаспор. Она редукционно делится с образованием четырех гаплоидных мегаспор, расположенных цепочкой друг над Другом по вертикали.
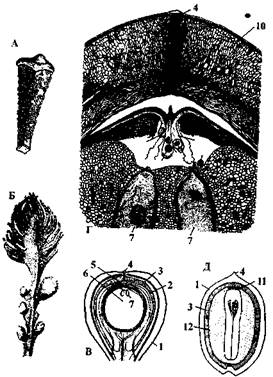
Рис. 42. Саговниковые (Cycadaceae): А — микроспорофилл цикаса (Cycas circinalis); Б — мегаспорофилл цикаса (С. revoluta); В — мегаспорангий (семязачаток) С. revoluta в продольном разрезе; Г — верхняя часть семязачатка С. revoluta в продольном разрезе (увеличено); Д — продольный разрез семени диоон (Dioon edule) (схема): 1 — наружный мясистый слой, 3 — внутренний мясистый слой, 3 — каменистый слой (склеренхима) интегумента, 4 — микропиле, 5 — пыльцевая камера, 6 — нуцеллус, 7 — архегонии, 8 — пыльцевые трубки, 9 — сперматозоиды, 10 — интегумент, 11 — корешок зародыша, 12 — эндосперм (женский гаметофит) Три верхние мегаспоры отмирают, а нижняя прорастает, образуя женский заросток — первичный эндосперм. Заросток, таким образом, находится внутри мегаспорангия семязачатка. На женском заростке формируется два архегония. Архегонии состоит из крупной яйцеклетки, рано отмирающего ядра брюшной канальцевой клетки и двух шей-ковых канальцевых клеток. Параллельно этому происходит неравномерное разрастание периферических клеток заростка над архегониями. В результате этого в центре формируется внутренняя полость, называемая архегониальной камерой. Постепенно дно пыльцевой камеры разрушается, и она сливается с архегониальной камерой. Микроспоры, прилипшие к капельке опылительной жидкости втягиваются в пыльцевую камеру нуцеллуса и прорастают. Экзина при этом лопается и через разрыв прорастает, растягивая интину, клетка гаустория. Она внедряется в стенку пыльцевой камеры и высасывает из ткани нуцеллуса питательные вещества. Генеративная клетка в это время делится на две, и одна из образовавшихся клеток — сперматогенная — начинает усиленно расти. В ней и формируются, не сразу, а спустя несколько месяцев, мужские гаметы-сперматозоиды. В большинстве случаев образуется два сперматозоида. Однако у некоторых видов образуется четыре, не менее 16 и до 22 сперматозоидов. Сперматозоиды саговников — самые крупные в растительном мире, достигают 3— 3,5 мм. К этому времени внутри пылинки создается высокое тургорное давление, так как гаустория активно поглощает влагу из клеток нуцеллуса. Это приводит к разрыву оболочки микроспоры. Слизь с заключенными в ней сперматозоидами выбрасывается на дно архегониальной камеры. Канальцевые клетки архегония к этому времени ослизняются, что облегчает доступ сперматозоидов к яйцеклетке. От опыления до оплодотворения иногда проходит 6—7 мес. Половой процесс не требует, в отличие от папоротников, свободной ка-пельно-жидкой воды. Он автономен благодаря специализированной гаустории, обеспечивающей создание влажной среды самим растением. У саговниковых сперматозоиды являются самыми крупными мужскими гаметами, известными науке. У замии флоридской их длина достигает 325 мкм и видны даже невооруженным глазом. После оплодотворения зигота делится и формирует зародыш, образуя подвесок, зачаточный стебелек, первичный корень, две семядоли и конус нарастания. Семязачаток превращается в семя. Интегумент становится покровом семени, эндосперм — кладовой питательных веществ для молодого спорофита (зародыша). В благоприятных условиях семя прорастает и дает начало новому растению. Цикл развития замии (Zamia) приведен на рис. 43. Необходимо отметить, что нуцеллус является новым образованием, которое отсутствовало у папоротниковидных. Саговниковые находят практическое применение. Из сердцевины получают крупу саго. Многие саговники — прекрасные декоративные растения.
Рис. 43. Цикл развития замии (Zamia): 1 — женское растение; 2 — мегаспорофилл с двумя семязачатками; 3 — образование линейной тетрады мегаспор; 4 — семязачаток с единственной сохранившейся мегаспорой; 5 — образование свободноядерного женского гаметофита; 6 — образование клеточного гаметофита; 7 — появление архегониев; 8 — мужское растение; 9 — микроспорофилл; 10 — микроспорангий; 11 — микроспора; 12 — образование трехклеточного мужского гаметофита в микроспоре; 13 — прорастание клетки — гаустории; 14 — образование сперматогенной клетки; 15 — формирование сперматозоидов; 16 — верхняя часть семязачатка перед оплодотворением; 17 — то же, после оплодотворения одной (левой) из яйцеклеток; 18 — проросток КЛАСС БЕННЕТТИТОВЫЕ {BENNE77ITOPSIDA) Вымершая группа. Существовала от нижнего триаса до верхнего мела (230—70 млн. лет назад). Наибольшую роль играли в юрском периоде и в верхнем мелу (195—70 млн. лет назад). Происходят, вероятно, от семенных папоротников. Листья перистые или реже цельные. Устьица сложногубые (синдетохейльные). Стебли маноксильные, как у саговниковых. Спорофиллы были собраны в стробилы обоеполые, реже однополые. Семязачатки с пыльцевой камерой. Интегумент семязачатка вытянут в длинную микропилярную трубку. Вымирание беннеттитовых совпало с возникновением цветковых растений. Беннеттитовые — небольшие деревья, имевшие облик саговников или папоротников. Стебли были или высокими, колонновидными, или обильно ветвились почти от основания. Листья у большинства были сложноперистыми, реже простыми, цельными. По строению стебля беннеттиты были сходны с саговниками.
Особенность беннеттитов — наличие обоеполой пазушной шишки-стробила (рис. 44). Снаружи шишка была одета верхушечными листьями, расположенными по спирали. Далее по оси шишки располагались кругами перистые (реже цельные) микроспорофиллы с микроспорангиями. Многочисленные микроспоры были без воздушных мешков. В верхней части шишки, заканчивая ось, располагались спарангиофо-ры в виде цилиндрических ножек с сидячими семязачатками на их вершинах. Они чередовались со стерильными чешуйками, которые на верхушке многоугольно расширялись, охватывая со всех сторон семязачаток и защищая его. Лишь в углах, сходившихся многоугольников оставались отверстия, через которые наружу выходили микропиляр-ные трубки, улавливающие пыльцу. Поэтому беннеттитов называют «полупокрытосеменными» растениями. Беннеттиты обладали уникальными среди голосеменных растений семенами — они не имели эндосперма. Их питательные вещества были сосредоточены в двух крупных семядолях, что считается прогрессивным признаком. После открытия беннеттитов их шишку (стробил) стали рассматривать как прототип цветка покрытосеменных, растений. Английские палеоботаники Э. Арбер и Дж. Паркин создали стробилярную гипотезу происхождения цветковых растений. В самом деле, обоеполая шишка беннеттитов сходна с цветком магнолии, но покрытосемянность у них другой природы, чем у цветковых растений. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|