
|
|
КЛАСС АНТОЦЕРОТОВЫЕ (ANTHOCEROTOPSIDA)
Свое название получили от греческих слов ап-thos— цветок, keros — рог. Спорогоны имеют вытянутую изогнутую в виде рога форму (рис. 9). Класс объединяет около 300 видов, шесть родов и два семейства. Это талломные растениям виде _^зеткоподобной_ „пл&^~. станки^ Чертой примитивной организации антоцеротовых являются пластинчатые хло-р"опл£ста1Гггйрёноидами, как у водорослей.
"* "Спорофит у антоцеротовых развит хорошо, имеет вид длинной коробочки, стопы и меристематического слоя между ними. Встречаются антоцеротовые в умеренных областях обоих полушарий и в тропиках. В странах СНГ растет два их вида: Anthoceros punc-tatum и Phaeoceros laevis. КЛАСС ПЕЧЕНОЧНИКИ, ИЛИ ПЕЧЕНОЧНЫЕ МХИ {MARCHANTIOPSIDA, HEPA7ICOPSIDA) Растения этого класса довольно разнообразны, являются таллом-ными. У одних печеночников гамедхфих=имеет вид слоевища — пластинки (сферокарповые, маршанциевые, большинство метцгериевых); у других вегетативное тело представлено листостебельным побегом (гапломитриевые, юнгерманиевые и некоторые метцгериевые). Ризоиды одноклеточные. Спорогон в виде коробочки на короткой ножке, хлорофилла не имеет. Со споры вырастает протонема (предросток) в виде небольшой пластинки, которая затем развивается во взрослый гаметофит. Класс включает около 8500 видов (280 родов) и подразделяется на два подкласса: маршанциевые (Marchantiidae) и юнгерманиевые (Jungermaniidae). Подкласс маршанциевых подразделяется на три порядка: маршанциевые {Marchantiales), сферокарповые (Sphaerocarpales) и моно-клиевые (Monocleales). Порядок маршанциевые (Marchantiales) Гаметофит этого порядка представлен талломом лентовидной формы, имеет дорзовентральное строение с внутренней дифференциацией и ризоидами двух видов. Спорофит в виде коробочки на короткой ножке, расширенная часть которой получила название гаустории или присоски. В коробочке имеются споры и элатеры (пружинки). Порядок объединяет 33 рода и 400 видов. Род маршанция насчитывает 65 видов. Наиболее известным представителем является маршанция обыкновенная (Marchantia polymorpha) (рис. 10). Она встречается на влажной почве в лесах на месте кострищ, по болотам, откосам осушительных канав и в других местах. Вид почти космополитный. Маршанция представляет собой лентовидный, дихотомически разветвленный, стелящийся по земле таллом 5—20 см длиной и 1— 2 см шириной. Верхушка таллома выемчатая с точкой роста. Верхняя сторона таллома темно-зеленая со средней ложбинкой; нижняя буровато-красная. Вдоль средней части таллома отходят многочисленные одноклеточные ризоиды. Здесь же хорошо заметны (в лупу) особые выросты таллома, чешуйки — амфигастрии — нижние однослойные брюшные чешуйки, которые тянутся в два ряда вдоль средней части, а также и вдоль краев таллома. Различают простые и язычковые ризоиды. Последние сплетаются в тяжи и тянутся вдоль краев таллома. Они прижимаются к таллому
Рис. 10. Маршанция обыкновенная {Marchantiapolymorpha): А — таллом с мужскими подставками; Б — таллом с женскими подставками; В — поперечный разрез таллома: воздушная камера с устьицем и ассимиляторами; Г — устьице с поверхности; Д — таллом с выводковыми корзинками; Е — разрез корзинки с выводковыми почками; Ж — выводковая почка; 3 — верхушка мужской подставки в разрезе, полости с антеридиями; И — антеридий: ножка, оболочка и сперматогенная ткань; К — группа архегониев на женской подставке; Л — архегоний, в брюшке его — яйцеклетка; М — спорогон: ножка, коробочка и остатки шейки архегония; Н — споры и одна пружинка, или элатера амфигастриями и образуют своеобразный фитиль (наружная проводящая система). Простые ризоиды прикрепляют таллом к субстрату и всасывают воду с растворенными минеральными веществами. Анатомическое строение таллома довольно простое. Он имеет верхнюю и нижнюю эпидерму, между которыми располагается ткань. Верхняя эпидерма имеет устьица. Под верхней эпидермой находятся 29 воздушные камеры — полости отграниченные крупными бесцветными клетками в 1—2 яруса. Со дна воздушных камер поднимаются ветвистые нити из клеток-ассимиляторов (с хлорофилловыми зернами). Под воздушными камерами располагается паренхиматическая ткань из широких бесцветных клеток, нередко содержащих крахмал и включения масла (масляные тела). В паренхиме можно обнаружить слизевые ходы, образованные в результате растворения соседних клеток, всегда заполненных слизью. От нижней эпидермы отходят ризоиды и амфигастрии. Последние прижимают к нижней стороне таллома маршанции язычковые ризоиды, которые, сплетаясь, образуют наружную проводящую систему, подобную на фитиль. Внутри таллома маршанции нет проводящей т-кани. Она заменяется наружной проводящей системой из язычковых ризоидов с амфигастриями. Масляные тела и слизевые ходы — характерные особенности всех печеночников. Вегетативное размножение маршанции происходит при помощи выводковых почек. На верхней стороне таллома маршанции имеются выводковые корзинки. В них формируются выводковые почки в виде овальной пластинки с выемками по бокам, где помещаются точки роста. Выводковая почка сидит на одноклеточной ножке. Во время дождя при попадании капли воды в выводковую корзинку выводковая почка легко отламывается от ножки и вымывается из нее. На поверхности почвы из выводковой почки формируется новый таллом маршанции. Таллом маршанции имеет дихотомическое ветвление. Он нарастает верхушкой и отмирает с противоположной стороны. Когда отмирание таллома доходит до разветвления, ветви растут самостоятельно. Для маршанции также характерно половое размножение. Маршанция — растение двудомное. На одних ее талломах возникают своеобразные выросты в виде ножки с многолопастным диском — мужские, антеридиальные подставки. На продольном разрезе через диск в верхней его части заметны антеридиальные полости, на дне которых расположены по одному антеридию — овальному тельцу с однослойной стенкой на ножке, которое прикрепляется ко дну полости. Содержимое антеридия состоит из многочисленных сперматогенных клеток, каждая из которых образует два двужгутиковых сперматозоида. После созревания антеридий вскрывается наверху щелью, сперматозоиды во время дождя через ка-
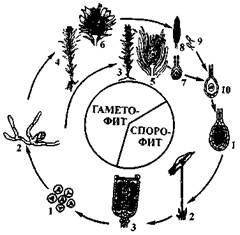
нал антеридиальной полости выходят наружу и, активно двигаясь в воде, подплывают к архегонию. На других талломах образуются выросты в виде ножки, заканчивающейся многолучевой звездой. Это женская, архегониальная, подставка. На продольном разрезе через женскую подставку видны архегонии, группами сидящие между лучами и обращенные шейками вниз. Каждая группа архегониев покрыта общим покровом — перихецием, а каждый архегонии одет частным покровом — перианцием в виде кольцевого валика у основания. Затем перианции сильно разрастается. Пе-рихеций и перианции защищают архегонии, а затем и спорогоны от высыхания. При созревании архегония шейка его на верхушке вскрывается, шейковые и брюшная канальцевые клетки ослизняются и сперматозоиды беспрепятственно проникают внутрь архегония. Происходит слияние сперматозоида с яйцеклеткой — оплодотворение. Из оплодотворенной яйцеклетки развивается бесполое поколение маршанции — спорофит, или спорогон. Он состоит из шаровидной коробочки, короткой ножки, расширенная часть которой называется гаусторией (ступней). При помощи гаустории спорогон внедряется в ткань подставки (гаметофита) и питается за его счет. Коробочка содержит споры и элатеры (пружинки). При созревании спор коробочка вскрывается на верхушке створками, и споры высыпаются наружу. Элатеры сильно гигроскопичны. В сырую погоду они скручиваются, в сухую — ракручиваются, разрыхляя споровую массу и способствуя равномерному высеиванию спор из коробочки. Споры гаплоидные, образуются при мейозе археспориалъных клеток (спороцит). Часть материнских клеток спор вытягивается, оболочка их неравномерно утолщается и принимает вид спиралек. Каждая такая клетка превращается в пружинку — элатеру. Спора одета двумя оболочками — наружной (экзоспорий) и внутренней {эндоспорий). Попав на почву, спора прорастает, образует пластинчатую протонему, которая затем формирует таллом маршанции. Цикл развития маршанции приведены на рис. 11. Таким образом, в цикле развития маршанции наблюдается чередование поколений (генераций), которым соответствует смена ядерных фаз. Половое поколение (гаметофит) сменяется бесполым (спорофитом).
Рис. 11. Цикл развития маршанции (Marchantiapolymorpha): 1 — мужской гаметофит, 2 — антеридий, 3 — сперматозоиды, 4 — женский гаметофит, 5 — архегонии, 6 — зигота, 7 — начальная стадия развития спорогона, 8 — спорогенная клетка, 9 —11 — развитие из спорогенной клетки четырех спор путем мейоза, 12 — раскрывшийся спорогон, 13 — прорастание споры, 14 — молодой таллом Преобладающим поколением является гаметофит — растение маршанции с мужскими и женскими половыми органами (антеридиями и архегониями). Все клетки его гаплоидны. Гаметофит берет начало от гаплоидной споры. Спорофит (спорогон) имеет небольшие размеры и питается за счет гаметофита, к которому прикрепляется при помощи гаустории. Все клетки спорофита диплоидны, так как развиваются из диплоидной зиготы. Переход от спорофита к гаметофиту происходит в результате редукционного деления при образовании спор.
Растения этого
окружены специальной открытой сверху оберткой. Спорогон имеет вид шарообразной коробочки на короткой ножке с гаусторией (рис. 12). Стенка коробочки спорогона однослойная, при созревании разрушается. Вместо элатер развиваются питательные клетки. Порядок вклю- чает два семейства: сферокарповые (Sphaerocarpaceae) и риелловые (Riellaceae). Порядок моноклеевые (Monocleales) Талломы моноклеевых очень крупные, без воздушных камер и устьиц. Имеются клетки с масляными тельцами. Антеридии погруженные, собраны в дисковидные образования. Спорогон на длинной ножке. Коробочка цилиндрическая, тонкостенная. Элатеры длинные. Представитель — моноклея (Monoclea). Подкласс юнгерманниевые (Jungermanniidae) В состав подкласса входит четыре порядка: метцгериевые (Metzgeriales), такакиевые (Takakiales), гапломитриевые {Haplomitria-les), юнгерманниевые {Jungermanniales), около 50 семейств, 250 родов и свыше 5000 видов. Это как талломные, так и листостебельные растения, чаще дорзовентрального строения, со слабой дифференциацией тканей. Порядок юнгерманниевые {Jungermanniales)
Это самый крупный по количеству видов порядок подкласса. Насчитывает свыше 40 семейств, более 200 родов и около 5000 видов. Распространены преимущественно в теплых странах южного, полушария, встречаются также и в странах СНГ. Среди них известны как талломные, так и листостебельные формы. Антеридии и архе-гонии имеют обычное для моховидных строение. Располагаются на верхней стороне тела, на особых веточках или в пазухах листьев. Коробочка спорогона внутри содержит споры и обычно пружинки, раскрывается четырьмя створками.
Пеллия {Pellia epi-phylla) (рис. 13) встречается в странах СНГ. Растет обычно по сырым лесным оврагам, возле ручьев и канав. Имеет небольшой пластинчатый, дихотомически разветвленный таллом с ризоидами. Дифференциация тканей слабая, органы размножения находятся на верхней стороне таллома. Растение однодомное. Из листостебельных юнгерманниевых достаточно интересны радула сплющенная {Radula complanata) и хилосцифус многопериантие-вый {Chyloscyphus polyanthus) (рис. 14). В Беларуси широко распространен первый вид, который встречается на стволах липы и дуба. Дорзовентральные побеги радулы длиной 2—4 см плотно прижаты к коре деревьев. Выделенное из дернинки растение имеет ветвистый стебель. Главный стебель и боковые веточки 3. Зак. 377
Рис. 14. Радула {Radula sp.) и хилосцифус (Chylosciphus sp.): А — радула: 1 — общий вид (на верхушке одной веточки виден колокольчатый перихеций), 2—антеридий и архегонии, 3 — молодой спорогон (н — ножка, г — гаустория, к — коробочка со спорами и пружинками), 4 — масляные тельца; Б — хилосцифус: 1 — общий вид, 2 — амфигастрия несут два ряда листьев. Каждый лист двулопастной. Спинная лопасть крупная, брюшная — мельче, загнута вниз и прижата к стеблю. Однослойные листья своими краями налегают друг на друга. В результате растение имеет дорзовентральное строение. Амфигастрии у радулы отсутствуют. Радула — однодомный печеночник. Архегонии группами располагаются на верхушке веточек, при этом точка роста идет на их образование и рост веточки прекращается. Архегонии имеют типичное строение с покрывалом — перихецием. Антеридии расположены на длинной ножке (см. рис. 14, А2). В них развиваются двужгутиковые сперматозоиды. После оплодотворения из зиготы развивается спорогон. Зрелый спорогон имеет коробочку, ножку 35 Из подкласса юнгерманниевых в Красную книгу Республики Беларусь внесены меркия ирландская (Moerckia hibernica), гимноколея вздутая (Gymnocolea inflata), скапания заостренная (Scapania apiculata), порелла плосколистная (Porella platyphylla). КЛАСС ЛИСТОСТЕБЕЛЬНЫЕ МХИ (BRYOPSIDA, ИЛИ MUSC1) Листостебельные мхи — самый крупный класс моховидных. Он включает 700 родов и 14500 видов, которые" встречаются "повсеме-стно — от полярных пустынь Арктики до льдов Антарктиды. Гаметрфит листостебельных мхов расчленен на стебель и листья. Стебель имеет радиальное строение, на нем тесной" сййралью располагаются листья. На__стебле__формируются органы полового размножения — антеридии и архегонии. "* Спорофит (спорогон) развивается из оплодотворенной яйцеклетки и состоит из коробочки, ножки и гаустории (стопы). В коробочке формируются только споры, элатеры отсутствуют. Из споры развивается протонема (предросток) в виде длинной разветвленной нити, реже она пластинчатая. В коробочке большинства мхов есть перистом (аппарат для регуляции рассеивания спор; его нет у сфагновых и андреевых мхов). Класс подразделяют на три подкласса: сфагновые (Sphagnidae), андреевые (Andreaeidae), бриевые (Bryidae). Подкласс сфагновые, или торфяные, мхи {Sphagnidae) Подкласс включает один порядок, одно семейство, один род и свыше 350 видов. Распространены сфагновые мхи в умеренных и холодных областях на равнинах северного полушария. Растут на болотах и в заболоченных лесах, образуют сплошные покровы. В горах тропиков не образуют больших зарослей. Произрастает сфагнум плотными дернинами. Отдельный стебель сфагнума имеет ветви трех видов: верхушечные веточки образуют компактную головку, средние — горизонтально отклонены и нижние — свисают вдоль стебля (рис. 15). Анатомическое строение растения довольно простое. Стебель в центре имеет сердцевину из паренхимных клеток, а за ней располагается древесинный цилиндр, окружающий сердцевину, из клеток, окрашенных в бурый цвет (придает стеблю прочность). Снаружи стебель покрыт эпидермисом (гиалодермисом) из мертвых пустых клеток.
Рис. 15. Мох сфагнум {Sphagnum sp.): А — общий вид; Б — строение листа: видны крупные водоносные клетки; В — часть поперечного разреза стебля: кора из крупных водоносных клеток; Г — поперечный разрез листа: хлорофиллоносные клетки чередуются с водоносными; Д — антеридии, сидящие на ножках между листьями; Е — верхушка с архегониями; Ж — продольный разрез споро- гона: коробочка на «ложной» ножке, неполная колонка, спорангий в виде колпачка, стенка коробочки с крышечкой на верхушке, остатки архегония; 3 — протонема с ризоидами его веса. Высокая гигроскопичность способствует быстрому заболачиванию тех мест, где сфагнум поселяется. Сфагновые мхи хорошие торфообразователи, поэтому их называют торфяными мхами. Роль ассимиляции выполняют листья верхушечных и торчащих ветвей. Свисающие ветви образуют вокруг стебля как бы чехол, фитиль, способствующий быстрому продвижению воды вдоль стебля. Стеблевые листья сфагнума по форме отличаются от веточных и имеют важное систематическое значение. Взрослые растения сфагнума лишены ризоидов. Дернина сфагнума ежегодно отмирает в нижней части и продолжает свой рост вверх. В условиях обводненности и недостатка кислорода отмершие части сфагнума не разлагаются до конца и образуют торф. При этом образуются органические кислоты, которые еще больше угнетают бактериальный процесс разложения торфа. В зависимости от климата и гидрологических условий скорость образования торфа колеблется от 0,22 до 2 мм в год. N Сфагновые мхи бывают однодомными и двудомными. В верхней части стеблей образуются антеридии и архегонии на разных побегах. Антеридии формируются на верхушечных веточках, имеют удлиненную форму буроватой окраски и заметны простым глазом. Антеридии вскрываются лопастями. В них образуется большое количество дву-жгутиковых сперматозоидов. Архегонии типичного строения образуются на боковых или некоторых торчащих веточках по 3—5 штук. Из оплодотворенной яйцеклетки формируется спорогон. Он состоит из шаровидной коробочки, которая ниже, после небольшой перетяжки, переходит в ножку. При помощи ножки (гаустории) спорогон внедряется в верхушку стебля. Ко времени созревания спорогона верхушка стебля удлиняется и выносит коробочку над покровными листьями. Эту часть стебля называют ложной ножкой. Сверху коробочка имеет крышечку. Внутри коробочки имеется приподнимающаяся со дна неполная колонка. Сверху колонки расположен куполообразный спорангий. В нем формируются редукционным путем споры. Клетки эпидермиса коробочки, кроме устьичных, содержат хлорофилл. Таким образом спорогоний сфагнумов питается отчасти самостоятельно. Ко времени созревания спор колонка и стенка спорангия разрушаются. Споры оказываются в полости коробочки. Затем крышечка сбрасывается с большой силой и споры выбрасываются наружу. На почве спора прорастает в однослойную пластинчатую протонему. На протонеме образуются ризоиды и почка, дающая олиственный побег. На территории Беларуси очень редко встречаются и внесены в Красную книгу сфагнум Линдберга {Sphagnum lindbergii) и сфагнум мягкий {Sph. molle). Подкласс андреевые мхи (Andreaeidae) Подкласс представлен горными растениями и включает один порядок Andreaeales, одно семейство, три рода и 92 вида. Стебель многократного ветвится (рис. 16), прикрепляется к субстрату ризоидами. Листья однослойные, иногда со средней жилкой.
Рис. 16. Андреевым мох (Andreaca rupestris): А — растение со спорангиями (слева—коробочка, которая вскрывается четырьмя трещинами); Б — верхушка стебля с антеридиями; В — архегоний; Г — спорогон (к — колонка, сп — спорангий со спорами, ар — остатки архегония) Антеридии и архегоний располагаются на верхушках разных ветвей. По строению и развитию половые органы напоминают таковые следующего подкласса Bryidae. Спорогон во многом напоминает аналогичный у сфагновых мхов: коробочка на ножке, колонка и куполообразный спорангий, наличие ложной ножки. Коробочка вскрывается четырьмя створками. Протонема пластинчатая. Мхи этого подкласса занимают промежуточное положение между сфагновыми и зелеными мхами. Со сфагновыми мхами их сближает строение листа (однослойность), строение спорогона, наличие ложной ножки. С зелеными мхами у них сходен внешний вид, строение и развитие антеридиев и архегониев. Из этого подкласса в Красную книгу Республики Беларусь внесена андрея скальная (Andreaea rupestris). Подкласс зеленые, или бурые, мхи {Bryidae) В подкласс зеленых мхов входит порядок Bryales, 85 семейств, около 700 родов и 14000 видов. Широко распространены, имеют ог- ромное значение в умеренных и холодных областях обоих полушарий. Несколько меньшее их в тропиках — встречаются главным образом в горах. Зеленые мхи принимают участие в образовании растительного покрова. На болотах они образуют сплошной покров и являются важными торфообразователями на низинных и переходных болотах. В хвойных лесах, на лугах, в горах, тундре покрывают почву на большом протяжении. Зеленые мхи характеризуются хорошо развитой нитевидной, разветвленной протонемой. В коробочке спорогона имеется перистом (специальный аппарат для равномерного и постепенного рассеивания спор). Один из представителей зеленых мхов — кукушкин лен обыкновенный (Polytrichum commune) (рис. 17).
Рис. 17. Кукушкин лен (Polytrichum commune): А — женское растение со спорогоном (с колпачком и без него); Б — мужское растение с розеткой на верхушке; В — поперечный разрез стебля: в центре — зачаточная стель; Г — поперечный разрез листа с ассимиляторами; Д — верхушка мужского растения с антеридиями и парафизами; Е — антеридий: ножка, оболочка и сперматогенная ткань; Ж — формы парафиз; 3 — верхушка женского растения: архегоний окружены листьями; И — архегоний: в шейке его-канальцевые клетки, в брюшке — яйцеклетка и над ней брюшная канальцевая клетка; К — протонема мха с ризоидами и почками; Л — продольный разрез коробочки: внутри спорангий вокруг колонки, наверху крышечка Распространен в хвойных лесах, на окраинах болот. Имеет простой, неветвистый, прямостоячий стебель длиной 30—40 см, густо покрытый линейно-шиловидными листьями. В нижней части стебля листья более мелкие, бурые. Ниже стебель становится бурым. В почве от него отходят многочисленные ржаво-бурые ризоиды. Стебель имеет сложное анатомическое строение. В середине стебля проходит концентрический проводящий пучок — зачаточная стель. В центре пучка находится ксилема из мертвых удлиненных клеток. Функционально соответствуют трахеидам более организованных высших растений. За ксилемой в один—два слоя расположено «крахмальное влагалище», Клетки которого заполнены крахмальными зернами. За ним расположено кольцо флоэмы. Пучок окружает кора. В ней видны листовые следы — ответвления от центральной стелы, идущие в листья. Снаружи стебель покрыт эпидермисом. Листья кукушкина льна имеют сложное строение. С нижней стороны они покрыты эпидермой из толстостенных клеток. Над эпидермой располагаются сильно утолщенные клетки механической ткани, а над ними — тонкостенная паренхима. От нее перпендикулярно вверх отходят однослойные пластинки, которые тянутся продольными рядами вдоль всего листа. Клетки этих пластинок содержат большое количество хлоропластов, их называют ассимиляторами. Расстояние между ассимиляторами микроскопическое. По принципу капиллярности вода из окружающей среды быстро заполняет эти пространства и прочно удерживается ассимиляторами. Таким образом, ассимиляторы играют еще и роль водоснабжения. При подсыхании происходит заворачивание листа на верхнюю сторону, он превращается в трубочку, в полости которой располагаются ассимиляторы, и находящаяся между ними влага не испаряется. По такому принципу построены листья у ряда других видов. В центре всех листьев зеленых мхов проходит жилка. Кукушкин лен — растение двудомное. На верхушке стеблей одних растений образуются антеридии, на других — архегонии. Они собраны группами и окружены верхушечными листьями. Антеридии имеют мешковидную форму на короткой ножке и покрыты однослойной стенкой. В антеридии образуется большое количество сперматогенных клеток. Каждая из них дает двужгутиковый сперматозоид. Антеридий вскрывается в дождливую погоду щелью на верхушке. Между антеридиями находятся парафизы — выросты стебля в виде однорядных нитей или расширенных на верхушке пластинок. На мужских растениях группа антеридиев окружена вегетативными листьями бурого цвета и имеет вид розетки. Точка роста стебля 41не идет на образование антеридиев, и стебель в дальнейшем прорастает образуя новый побег. На стебле кукушкина льна можно видеть нередко 5—6 таких розеток. Архегонии имеют обычное строение, состоят из брюшка и длинной шейки. Снаружи они окружены листьями, не отличающимися от обычных вегетативных листьев. После оплодотворения из зиготы развивается спорогон. Он состоит из коробочки и длинной ножки, нижняя расширенная часть которой называется гаусторией. При помощи гаустории молодой спорофит получает питательные вещества от гаметофита. Сверху коробочка спо-рогона покрыта колпачком, или калиптрой (остатком брюшка архего-ния), впоследствии сбрасывается. Зрелая коробочка имеет сложное строение. Средняя расширенная часть ее получила название урночки, у основания коробочки — апофиза (шейка) и сверху — крышечка. Внутри коробочки от основания вверх поднимается колонка — тяж бесплодных паренхимных клеток. Колонка в верхней части расширяется и образует на границе крышечки и урночки тонкую пластинку — эпифрагму. Вокруг колонки подвешен на тонких нитях спорангий. Нити прикрепляют спорангий к колонке и стенкам урночки. В спорангии из спо-рогенной ткани путем редукционного деления образуются многочисленные гаплоидные споры. Окраина вскрывшейся коробочки имеет перистом. Он может иметь вид зубчиков, ресничек, сеточки. Перистом гигроскопичен. Во влажную погоду он набухает и занимает горизонтальное положение, стягивая к центру края коробочки. Зубчики перистома налегают на эпифрагму и закрывают коробочку целиком. Следовательно, перистом препятствует попаданию влаги внутрь коробочки и предотвращает слипание и загнивание спор. В сухую погоду зубчики перистома отгибаются наружу и не препятствуют высеиванию спор. Клетки стенки коробочки содержат хлорофилловые зерна. Поэтому спорогон зеленых мхов питается отчасти самостоятельно. Перистом — сложный орган, обеспечивающий равномерное и постепенное рассеивание спор. На поверхности почвы спора прорастает в нитчатую, обильно ветвящуюся протонему (предросток). На протонеме образуются почки, развивающиеся в листостебельные растения. На этом цикл развития кукушкина льна (рис. 18) заканчивается. Здесь, как и у всех зеленых мхов, преобладающим поколением является гаметофит — протонема, само листостебельное растение с антеридиями и архегониями. Спорофит представлен коробочкой на ножке, он 4. Зак. 377
тесно связан с гаметофитом, не может существовать самостоятельно. При образовании спор происходит редукционное деление. Споры, таким образом, гаплоидны. Вегетативное размножение осуществляется участками протонемы, образованием вторичной протонемы, выводковыми почками на листьях и стеблях.
По форме роста зеленые мхи подразделяют на верхоплодные и бокоплодные мхи. У верхоплодных (Асго-carpi) фхегонии, а затем спо-рогоны развиваются на верхушке главного стебля; у бокоплодных {Pleurocarpi) — на верхушке боковых ветвей. Представителями верхоплодных мхов являются Polytrichum commune, Funaria hygrometrica, Atrichum undula-tum, Bryum ventricosum, B. argenteum, Mnium undula-tum, Dicranum scoparium и др.; бокоплодные мхи: Pleurozium schreberi, Hylocomium proliferum, Ptilium crista-castrensis, Drepanocladus aduncus, Calliergon cordifolium и др. На территории Республики Беларусь редко встречаются и внесены в Красную книгу следующие представители подкласса бриевых мхов: цинклидотус дунайский (Cinclidotus danubicus), бриум Клин-греффа (Bryum klinggraeffii), птеригинандрум нитевидный {Pteriginan-drumfiliforme), паралевкобриум длиннолистный (Paraleucobryum longi-folium), тортелла извилистая (Tortella tortuosa), цинклидиум стигийский {Cinclidium stygium), ортотрихум Лайеля (Orthotrichum Lyellii), ринхостегиум стенной (Rhynchostegium murale). У ботаников нет единого мнения о происхождении и эволюции моховидных. Это древняя группа наземных растений. Достоверные остатки вегетативных органов моховидных известны из верхнекаменноугольных отложений как талломных, так и листостебельных мхов. Споры моховидных в последнее время находят в отложениях силура и девона. Современные палеоботанические материалы доказывают, что моховидные — очень древняя и обособленная группа высших растений. Классы Anthocerotopsida, Marchantiopsida и Btyopsida обособились очень давно, возможно, в силуре или раньше. Талломные формы появились раньше листостебельных. Сфагновые мхи представляют очень древнюю группу растений палеозойского возраста (см. табл. 2). Некоторые ученые считают, что моховидные берут начало от древних водорослей, которые имели стелющийся по субстрату таллом. Цикл развития их характеризовался изоморфным чередованием поколений. Они имели многоклеточные гаметангии и спорангии, как у современных бурых водорослей. В условиях наземного обитания их многоклеточные гаметангии в процессе эволюции дали начало антеридиям и архегониям, спорангии — спорофиту. В последнее время все более распространяется риниофитная гипотеза происхождения моховидных. Еще Р. Кидстон и У. Ланг высказали предположение, что моховидные произошли от риниофитов в результате общей редукции спорофита и прогрессивного развития гаметофита. Эту гипотезу поддержали ряд выдающихся ботаников: британский палеоботаник Д. Скотт, шведский палеоботаник Т. Халле, русский ботаник Б.М. Козо-Полянский и др. В пользу этой гипотезы свидеьельстуют и некоторые палеоботанические данные. У хорнеофи-тона (Horneophyton) из риниофитов в спорангии имеется колонка, подобная на колонку в коробочке сфагнума и андрея. Девонский род Sporogonites имеет спорофит с виде длинной ножки со спорангием, у него имеется колонка, как у сфагновых мхов. Американский палеоботаник Г. Эндрюс также сближает Sporogonites с моховидными. Наличие устьиц, на спорофите моховидных является весомым доказательством происхождения мхов от растений с хорошо развитым спорофитом. Такими растениями могли быть, скорее всего, ри-ниофиты. Значение моховидных заключается в том, что печеночники и зеленые мхи являются пионерами зарастания субстратов высшими растениями.
Процесс торфообразования на болотах связан с мхами, особенно сфагновыми. Торф имеет важное значение как топливо, сырье для химической промышленности, удобрение и изоляционный материал. При сухой перегонке торфа получают ценные продукты: воск, парафин, фенол, карболовую кислоту, аммиак, уксусную кислоту, метиловый спирт, краски, сахарин и др. При гидролизе 1 т сухого сфагнового торфа получают 120 л спирта. По самым скромным оценкам торфяники покрывают не менее 1% земной суши. Мировые запасы торфа составляют свыше 260 млрд. т. По запасам торфа страны СНГ занимают первое место в мире. В Беларуси торфяники занимают площадь более 2,5 млн. га с оставшимся запасом торфа в 2,3 млрд. т. Но в результате осушения болот возникает вопрос о рациональном использовании торфа, охране болот. В медицине торф применяют в торфотерапии. Известны курорты в Тверской, Полтавской областях. В годы Великой отечественной войны 1941—1945 гг. сфагнум использовался как заменитель ваты. Вопросы и задания для самоконтроля 1. Охарактеризуйте отдел моховидных. 2. Назовите черты примитивной организации печеночных мхов. 3. Дайте характеристику сфагновых мхов. 4. Что свидетельствует о промежуточном положении андреевых мхов между сфагновыми и бурыми мхами? 5. Чем подтверждается более высокая организация бурых мхов? 6. Как объясняется происхождение мхов? ОТДЕЛ ПЛАУНОВИДНЫЕ {LYCOPODIOPHYTA) Почти достоверно предками плауновидных можно считать зосте-рофиллофитов. Наибольшего развития плауновидные достигли в позднем палеозое (см. табл. 2). В настоящее время этот отдел представлен небольшим числом родов и видов. Современные представители — многолетние травянистые растения, обТЗчнсГ вечнозеленые: 'Среди ис-копа!шых~"Т5ыЛИ как~^трЗШЙШ^шё^"так~я"моТц11ыё"др^весные формы. Плауновидные представлены как равноспоровыми, так и разноспоро-выми растениями. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|




 Порядок сферокарповые {Sphaerocarpales)
Порядок сферокарповые {Sphaerocarpales)






 44
44