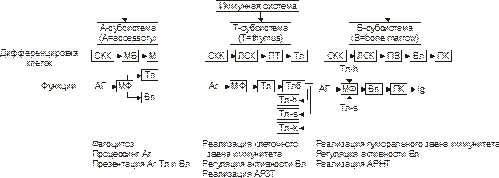|
|
Иммунокомпетентные клеткиК иммунокомпетентным клеткам относят T- и B-лимфоциты, а также NK-клетки и антигенпредставляющие клетки (рис. 17–3). Ы ВЁРСТКА Вставить файл «ПФ Рис 17 03Структура иммунной системы»
Рис.17–3.Организация и функции системы иммунокомпетентных клеток. СКК — стволовая кроветворная клетка, МБ —миелобласт, М —миелоцит, ЛСК — лимфопоэтическая клетка, ПТ — преT-лимфоцит, Тл — T-лимфоцит, Вл — B-лимфоцит, ПК — плазматическая клетка, Мф — макрофаг, Тлб — T-лимфоцит в состоянии бласттранcформации, Тл‑h — T-лимфоцит‑хелпер, Тл‑s — T-лимфоцит‑супрессор, Тл‑k — T-лимфоцит‑киллер, АРЗТ — аллергическая реакция замедленного типа, АРНТ — аллергическая реакция немедленного типа. B-лимфоциты Субсистема B-лимфоцитов образована различными их клонами. Название субсистемы отражает то обстоятельство, что лимфоциты, представляющие её, формируются у птиц в сумке (bursa) Фабрициуса (впервые B-лимфоциты были выявлены в лимфоидных органах птиц). У человека подобной бурсы нет. B-лимфоциты созревают в костном мозге, а также, возможно, в пейеровых бляшках, миндалинах, определённых зонах селезёнки и лимфоузлов. B-лимфоциты берут начало от стволовых кроветворных клеток костного мозга. B-лимфоциты обеспечивают реализацию эффекторного звена гуморального иммунного ответа. В мембране B-лимфоцита имеется рецептор для антигенов: мономер IgM. Из красного костного мозга B-лимфоциты мигрируют в тимус-независимые зоны лимфоидных органов. Продолжительность жизни большинства B-лимфоцитов не превышает десяти дней, если они не активируются Аг. Зрелые В-лимфоциты (плазматические клетки) вырабатывают иммуноглобулины всех известных классов. Для идентификации B-клеток выявляют их основные маркёры: CD19, CD20 и CD22. В процессе формирования B-клеток выделяют антигеннезависимую и антигензависимую стадии. Антигеннезависимая стадия созревания B-лимфоцитов происходит под контролем локальных клеточных и гуморальных сигналов от микроокружения пре‑B-лимфоцитов и не определяется контактом с Аг. На этой стадии происходит формирование отдельных пулов генов, кодирующих синтез Ig, а также экспрессия этих генов. Однако, на цитолемме пре‑B-клеток ещё нет поверхностных рецепторов: иммуноглобулинов. Компоненты их находятся еще в цитоплазме. Образование B-лимфоцитов из пре‑B-лимфоцитов сопровождается появлением на их поверхности первичных иммуноглобулинов, способных взаимодействовать с Аг. Только на этом этапе B-лимфоциты попадают в кровоток и заселяют периферические лимфоидные органы. Сформировавшиеся молодые B-клетки накапливаются в основном в селезёнке, а более зрелые — в лимфатических узлах. Антигензависимая стадия развития B-лимфоцитов начинается с момента контакта этих клеток с Аг (в том числе — аллергеном). В результате происходит активация B-лимфоцитов, протекающая в два этапа: пролиферации и дифференцировки. Пролиферация B-лимфоцитов обеспечивает два важных процесса:1) увеличение числа клеток, дифференцирующихся в плазматические клетки, продуцирующие АТ (Ig). Каждая плазматическая клетка способна секретировать несколько тысяч молекул в секунду. Процессы деления и специализации B-клетки осуществляются не только под влиянием Аг, но и при обязательном участии T-лимфоцитов‑хелперов, а также выделяемых ими и фагоцитами цитокинов: факторов роста и дифференцировки; 2) образование В-лимфоцитов иммунологической памяти. Эти клоны B-клеток представляют собой долгоживущие рециркулирующие малые лимфоциты. Они не превращаются в плазматические клетки, но сохраняют иммунную «память» об Аг. Клетки памяти активируются при повторной их стимуляции тем же самым Аг. В этом случае B-лимфоциты памяти (при обязательном участии T-клеток‑хелперов и ряда других факторов) обеспечивают быстрый синтез большого количества специфических АТ, взаимодействующих с чужеродным Аг, и развитие эффективного иммунного ответа или аллергической реакции. T-лимфоциты Субсистема T-лимфоцитов представлена различными их клонами. Т‑лимфоциты реализуют клеточное звено иммунного ответа, а также участвуют в регуляции его гуморального звена. Пролиферация и дифференцировка Т-лимфоцитов происходит под контролем вилочковой железы. В связи с этим их обозначают как T-клетки, или тимусзависимые лимфоциты. T-лимфоциты, как и B-лимфоциты, развиваются из стволовых кроветворных клеток костного мозга. Отсюда в виде клеток–предшественниц T-лимфоциты попадают с кровью в тимус, где происходит их антигеннезависимое созревание, сопровождающееся экспрессией на цитолемме специфических (у каждого лимфоцита своего) рецепторов. Т-клетки состоят из функциональных подтипов: CD4+ и CD8+. CD4+ Т-клетки или Т‑хелперы (TH) при их активации синтезируют и секретируют цитокины (ИЛ2, ИЛ4, ИЛ5, ИЛ6, γ-ИФН). В ходе иммунного ответа Т-хелперы взаимодействуют с молекулами MHC класса II. Цитотоксические T-лимфоциты (TC) или CD8+ Т-клетки, уничтожают инфицированные вирусом, опухолевые и чужеродные клетки при участии цитолитического белка — перфорина. CD8+ Т-клетки взаимодействуют с молекулой MHC класса I плазматической мембраны клетки–мишени. T‑супрессоры (TS) — представители CD8+ Т-клеток — регулируют интенсивность иммунного ответа, подавляя активность TH клеток; предотвращают развитие аутоагрессивных иммунных реакций и чрезмерного воспаления. NK-клетки NK-клетки (МНС‑нерестригированные киллеры, естественные киллеры) составляют до 15% всех лимфоцитов крови. Они не имеют поверхностных детерминант, характерных для T- и B-лимфоцитов, а также не имеют рецептора Т-лимфоцитов. В типичных NK-клетках экспрессируются дифференцировочные Аг CD2, СD7, CD56 и CD16 (рецептор Fc‑фрагмента IgG). В плазматической мембране активированных NK-клеток появляется гликопротеин CD69. NK-клетки распознают и уничтожают опухолевые и вирус-инфицированные клетки. Механизм распознавания неясен. Существует представление о наличии поверхностноклеточных молекул, защищающих клетки организма от цитотоксического действия NK-клеток. Примером служит продукт гена HLA‑C. Распознавание рецептором NK-клетки этой молекулы тормозит цитотоксическую активность NK-клеток и таким образом защищает клетку, экспрессирующую HLA‑C. Модификация продукта гена HLA‑C вирусами или связанными с опухолью молекулами приводит к уничтожению ее NK-клеткой. NK-клетки, располагая рецептором IgG (CD16), способны также взаимодействовать с клетками, окружёнными молекулами IgG, и уничтожать их (феномен АТ‑зависимой цитотоксичности). Активированные NK-клетки выделяют Цитолиз. В отличие от цитотоксических T‑лимфоцитов, способность NK-клеток к цитолизу не связана с необходимостью распознавания молекул MHC на поверхности мишени. NK-клетки уничтожают клетку–мишень не путём фагоцитоза, а (после установления с ней прямого контакта) при помощи перфорина. Гуморальная регуляция. Активность NK-клеток регулируется цитокинами. γ-ИФН и ИЛ2 усиливают цитолитическую активность NK-клеток. NK-клетки, наряду с макрофагами, нейтрофилами и эозинофилами, участвуют также и в АТ-зависимом клеточно-опосредованном цитолизе. Для этого NK-клетки экспрессируют на своей поверхности рецептор Fc‑фрагмента IgG (CD16). Fc‑фрагмент этих АТ взаимодействует с рецептором Fc‑фрагмента, встроенным в плазматическую мембрану NK-клетки. Антигенпредставляющие клетки (АПК илиА-субсистема) (рис. 17-3) присутствуют преимущественно в коже, лимфатических узлах, селезёнке и тимусе. К АПК относят макрофаги, дендритные клетки, фолликулярные отростчатые клетки лимфоузлов и селезёнки, клетки Лангерханса, М‑клетки в лимфатических фолликулах пищеварительного тракта, эпителиальные клетки вилочковой железы. Эти клетки захватывают, перерабатывают и представляют Аг (эпитоп) на своей поверхности другим иммунокомпетентным клеткам, вырабатывают ИЛ1 и другие цитокины, секретируют простагландин E2 (PGE2), угнетающий иммунный ответ. Фагоцитарную и цитолитическую активность макрофагов усиливает Дендритные клетки (ДК) образуются в костном мозге и формируют популяцию долгоживущих клеток, которые запускают и модулируют иммунный ответ. В костном мозге их предшественники образуют субпопуляцию CD34+-клеток, которые способны дифференцироваться в клетки Лангерханса для эпителия и дендритные клетки для внутренней среды. Незрелые и неделящиеся предшественники ДК заселяют многие ткани и органы. Дифференцировку ДК поддерживают колониестимулирующий фактор гранулоцитов и макрофагов GM‑CSF и ИЛ3. ДК имеют звёздчатую форму и в состоянии покоя несут на поверхности относительно небольшое количество молекул МНС. В отличие от клеток Лангерханса, интерстициальные ДК способны стимулировать синтез Ig В‑лимфоцитами. Все ДК могут вначале поступать в тимус-зависимую зону периферических лимфоидных органов, где созревают в так называемые интердигитирующие клетки. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|