|
|
Какой рецептор обеспечивает миграцию Т-лимфоцитов и дендритных клеток в Т-зоны лимфоидных органов?● CCR7] 31. Физиологический смысл положительной и отрицательной селекции тимоцитов? 1) Положительная селекция = Удаление клеток, не способных распознавать пептиды в составе молекул MHC.Если у TCR DP-тимоцита есть сродство к MHC (расположенному на кортикальных стромальных клетках тимуса)- он получает сигнал выживания, вызывающий увеличение экспрессии Bcl-2 (антиапоптотический фактор). Получение CD4+CD8+ T-клетками, (чувствительными к апоптозу) сигнала выживания. 90% клеток не проходят +селекции в тимусе и погибают в результате митохондриального апоптоза. 2) Отрицательная селекция - Выбраковка потенциально аутоагрессивных Т-клеток - обладающих слишком высоким сродством к МНС, следующий этап. Они получают сигнал к апоптозу. Гибнет порядка 50% тимоцитов, прошедших положительную селекцию. 32. Роль пролиферации лимфоцитов в развитии иммунного ответа. Исходная численность клеток в каждом клоне лимфоцитов мала и недостаточна для защиты организма от патогенов и других источников биологической агрессии. Например, пролиферация обеспечивает увеличение численности цитотоксических Т-клеток в 50 000 раз, чего достаточно для реализации их эффекторной функции.
33. Значение апоптоза в развитии лимфоцитов и в иммунном ответе. 1) В развитии лимфоцитов: ● Процесс апоптоза заложен в основу положительной и отрицательной селекции В и T-лимфоцитов ● Формирование популяций лимфоцитов (гибель избыточных клеток, которым не хватило гомеостатических факторов) ● Формирование антиген-распознающих рецепторов (Выбраковка клеток с дефектами реаранжировки рецепторных генов) ● Дифференцировка субпопуляций (Апоптоз Т-клеток при несоответствии специфичности рецептора и корецептора) 2) В зрелом периоде ● Гомеостатический контроль численности зрелых клеток ● Элиминация старых клеток 3) Иммунный ответ ● Активация и пролиферация лимфоцитов (Активационный апоптоз - передача сигнала через мембранные рецепторы) ● Созревание аффинности антител (Гибель низкоаффинных клонов В-клеток) ● Элиминация эффекторных клеток (Апоптоз отработавших клеток[2] ) ● Реализация цитотоксического эффекта лимфоцитов (Гибель клеток-мишеней цитотоксических лимфоцитов) 34. Какие каспазы и в какой последовательности активируются при запуске апоптоза Fas рецептором? Fas-ligand+Fas-R→FADD(Fas-Associated protein with Death Domain - белок с доменом смерти) →активация каспазы 8 (точнее прокаспазы 8) (тоже есть домены смерти, через них она связывается с FADD)→активация каспазы 3→активация каспаз 6 и 7
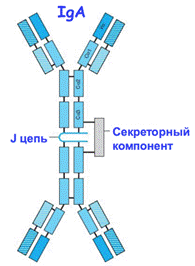
35. Особенности распознавания антигенов рецепторами В- и Т-клеток. B-клетки способны распознать нативный антиген, поскольку их рецептор это и есть антитело, а αβТ-клетки распознают антиген только в контексте МНС. 36. Схема строения молекулы антитела. Изотипы антител. 1) константная часть (эффекторные функции, опосредуется Fc рецепторами) и 2) вариабельный домен (Fab), распознавание антигена. 2 тяжелые (Н) и 2 легкие (L) цепи, дисульфидная связь Классы антител: IgG – иммуноглобулин крови. Долгосрочный иммунитет, нейтрализация токсинов, проникновение через плаценту, активация комплемента, активация фагоцитов. Подклассы IgG1, IgG2, IgG3, IgG4. IgM – первая линия защиты, активированные В лимфоциты производят эти АТ первыми. IgA – секретируемое АТ, защита на слизистых поверхностях. Подклассы IgA1, IgA2. IgE – нейтрализация паразитов, аллергические реакции. IgD – функция точно не известна, есть в наивных В лимфоцитах, необходим для корректной активации В-клеток. Все 5 классов антител представлены у всех животных, имеющих антитела, но различаются по аминокислотному составу и количеству доменов. IgG/IgD/IgE - мономер IgM - пентамер c 10ю антиген-связывающими участками -> увеличение авидности (стабильности комплекса антиген-антитело): мономерный IgM обладает низкой аффинностью. IgA - димер с секреторным компонентом и J-цепью.
37. Строение вариабельных доменов антител. Вариабельные домены легкой и тяжелой цепи содержат по 4 каркасных участка (framework region) и 3 гипервариабельных (hypervariable region; complementarity determining region). Гипервариабельные домены легкой и тяжелой цепи формируют антигенсвязывающий участок антитела. Именно сверхвысокая вариабельность этих участков обеспечивает возможность узнавания огромного числа антигенов. (Катя Кропочева, физраст, из Ярилина и лекций). 38. Физиологический смысл V(D)J перестройки генов иммуноглобулинов. Генерация разнообразия антиген-распознающих структур лимфоцитов - Ярилин. 39. Функция белков RAG1 и RAG2. ● Эндонуклеазы - отвечают за включение процесса реаранжировки V-генов ● Распознают и связываются с RSS (консервативные сигнальные последовательности, правило 23/12 в тяжелой цепи и 12/23 в легкой) и сближают их вместе. Затем инициируют двухцепочечный разрыв ДНК и делают шпильку - негомологичная рекомбинация. 40. Функция белка TdT в перестройке генов рецепторов лимфоцитов. фермент терминальная дезоксинуклеотидилтрансфераза обеспечивает нематричное добавление нуклеотидов при D-J рекомбинации - формирование N-вставок
41. Функции мембранной и секреторной форм BCR и переключение между ними. На В-клетках представлены В-клеточные рецепторы(BCR — B-cell reseptors), имеющие иммуноглобулиновую природу. При дифференцировке В-лимфоцитов в плазматические клетки в ходе иммунного ответа эти рецепторы секретируются в растворимой форме, называемой антителами. BCR распознают свободный и связанный с мембраной антиген, точнее, фрагмент молекулы антигена, называемый эпитопом. С В-клеточного рецептора начинается цепь передачи сигнала внутрь клетки, которая в зависимости от условий может приводить к активации, пролиферации, дифференцировке или апоптозу В-лимфоцитов. Известно, что В-клеточный рецептор играет особую роль в развитии и поддержании злокачественных В-клеточных заболеваний крови. Функции секреторных форм иммуноглобулинов: 1. нейтрализация токсинов/бактерий 2. опсонизация, стимулирующая фагоцитоз 3. опсонизация, стимулирующая антителозависимую цитотоксичность 4. комплемент-зависимый цитолиз Переключение происходит при дифференцировке активированных B-лимфоцитов в антитело-образующие клетки - плазмоциты. Механизм заключается в разном процессинге - на 3’ конце С-гена polyA ставится за SC сегментом, а не за MC. MC даёт гидрофобность, SC наоборот - повышенную растворимость. Регуляция переключения между изоформами происходит на уровне альтернативного сплайсинга. Лимфоцит может одновременно делать разные продукты: секретируемые Ат и BCR.
42. Физические силы, участвующие во взаимодействии антигена с антителом. Аффинность взаимодействия антигена и антитела. Эпитоп - участок антигена, непосредственно взаимодействующий с антителом; антиген-распознающий домен антитела необязательно непрерывная последовательность - это могут быть несколько пространственно разнесённых участков (см. CDR-домены в 37). Между эпитопом антигена и антителом устанавливаются связи: - Электростатические - Водородные - Гидрофобные - Ван-дер-Ваальса. +необходимо пространственное соответствие эпитопа и акт. центра антитела, т.к. эти связи действуют на не очень большом расстоянии -> аффинность антитела к антигену привязана к пространственной конфигурации и нарушается при её изменении. . Аффинность - мера сродства антител к антигену и может рассматриваться как показатель специфичности антител к данному эпитопу.
43. Селекция В-лимфоцитов. Роль клеточной гибели, индукции анергии. Редактирование V-генов. ● Всё в костном мозге! ● Под действием факторов EBF, E2A, Pax5 клетка-предшественник начинает перестройку V-генов и становится про-В-клеткой (progenitor). ● Про-B-клетка имеет перестроенные гены тяжелых цепей. Для продолжения развития ей необходимо пройти стадию селекции. Про-В секретирует пре-BCR, который состоит из 2х перестроенных тяжелых цепей и 2х инвариантных предшественников легких цепей. Сборка такого комплекса (лиганд не нужен!) сигнализирует об успешной перестройке, и клетка переходит в стадию пре-В-клетки (precursor). Отсутствие сборки комплекса -> смерть. Клетка становится чувствительна к IL-7, который стимулирует её пролиферацию. Также сборка пре-BCR включает механизм аллельного исключения - блокировку перестройки генов тяжелых цепей второй хромосомы. Происходит перестройка легких цепей, так же включается аллельное исключение. Перед покиданием костного мозга пре-B-клетка экспрессирует IgM на мембране. Происходит тестирование на аутореактивность. Аутореактивные клетки -> 1) удаление клонов (апоптоз); 2) анергия (неспособность отвечать на лиганды); 3) receptor editing - переарранжировка вторых аллелей
44. Способы участия антител в иммунной защите. 1)Нейтрализация, 2)Опсонизация, 3)Активация комплемента, 4)Антитело-зависимая клеточная цитотоксичность (ADCC).
45. Эффекторные функции антител различных изотипов антител. Классы антител: IgG – им/гл крови. Долгосрочный иммунитет, нейтрализация токсинов, проникновение через плаценту, активация комплемента, активация фагоцитов. IgM – первая линия защиты, активированный ВЛц производит эти АТ первыми. IgA – секретируемое АТ, защита на слизистых поверхностях, особенно, в кишечнике IgE – нейтрализация паразитов, аллергические реакции. IgD – функция точно не известна, есть в наивных ВЛц, необходим для корректной активации В-клеток.
46. Роль Fc–рецепторов в гуморальном иммунном ответе 1) опсонизация патогенов к поглощению их фагоцитами - Fc-рецепторы на поверхности фагоцитов запускают фагоцитоз, связываясь с антителами на поверхности фагоцитируемой частицы (это конечно уже клеточный ответ, но Fc рецепторы осуществляют связь между гуморальным и клеточным ответами) (см. вопрос 18) 2) антитело-зависимая цитотоксичность NK-клеток, Fc рецепторы на поверхности NK-клеток запускают киллинг после связывания с антителами (не совсем гуморально, но Купраш считает, что надо упомянуть) 3) участие в захвате иммунных комплексов фолликулярными дендритными клетками, которые сидят в лимфоузлах и участвуют в процессе созревания В-клеток - увеличения их аффинности (способности секретируемых ими антител связывать антиген) и переключение изотипов антител 47. Особенности строения и функции секреторного IgA. Защита на слизистых поверхностях. Секреторный IgA связывается с поверхностью патогенов, блокируя их адгезию на слизистых оболочках и подвижность. IgA - димер. Два мономера скреплены своими С-концевыми участками специальным полипептидом - J-цепью (он же участвует в олигомеризации IgM) и секреторным компонентом SP, обеспечивающим транспорт через эпителиальные слои и защищающим Ig A от протеолиза.
48. Свойства и локализация плазматических клеток. Плазматические клетки -активированные В-клетки, непосредственно секретирующие антитела. Свойства: ● секретируют Ig любого класса. ● крупные, развитый ЭПР ● живут 4-7 недель, в костном мозгу - десятки лет (аж до 75 лет!) ● полностью неподвижны и не отвечают на внешние стимулы ● маркер - CD138 Локализация: костный мозг, красная пульпа селезенки
49. Созревание аффинности и переключение изотипов антител при иммунном ответе. Ограниченное содержание антигена в зародышевых центрах вызывает конкуренцию за его связывание между антигенспецифичными В-клетками, в которой побеждают В-клетки, несущие BCR, обладающие наибольшим сродством к антигену. Этот процесс называют созреванием аффинитетаантигенраспознающих рецепторов (или антител, поскольку мембранные рецепторы на следующих этапах дифференцировки В-клеток будут секретироваться в виде растворимых иммуноглобулинов — антител). Параллельно происходит переключение изотипа секретируемых иммуноглобулинов с IgM на IgG, IgA и IgE. 50. Ig каких изотипов присутствуют на поверхности зрелых наивных B-клеток? M, D.
51. Каковы сигналы помощи В клеткам со стороны Т клеток ? Главное: ● TCR/MHCII -> Tfh (фолликулярные Т-кл) цитокины -> действуют на В-клетку ● B7/CD28 -> Tfh цитокины ● CD40/CD154 Факторы для пролиферации B-клеток: IL-4, IL-2, IL-5. 52. Какой физиологический смысл миграции В клеток в зародышевые центры? ● Созревание аффинитета и переключение изотипов (M→G, A, E) ● В-клетки, мигрируя в зародышевые центры, проходит путь Т-зависимой дифференцировки с участием фолликулярных дендритных клеток и CD4+-фолликулярных Т-хелперов. В этих В-клетках происходит переключение изотипов и созревание аффинитета. Они дифференцируются в долгоживующие антителообразующие клетки. То есть физиологический смысл этого процесса – образование плазмоцитов и B-клеток зародышевого центра и последующая продукция антител и В-клеток памяти для формирования иммунологической памяти. 53. Гибридомы и моноклональные антитела. Области применения. Гибридомы являются бессмертными клеточными клонами, продуцирующие антитела одной специфичности. Гибридомы получают при слиянии нормальных лимфоцитов , продуцирующих антитела, с подходящей линией плазмацитом (опухоли В-клеток, грубо говоря) с последующей селекцией на селективной среде. совмещают 3 свойства: 1. способность к неограниченному росту, 2. иммуноспецифичность - образуют моноклональные антитела, абсолютно идентичные 3. высокий уровень продукции АТ. Использование: ● терапевтические средства (например, против меланомы, рака груди и других видов рака и аутоимунных заболеваний - рассеянный склероз, ревматоидный артрит и т.д.) ● разные тесты (ELISA, FACS, western blot и т.д.)
54. Основные механизмы и проявления различных вариантов аллергии. ● 1-й тип — гиперчувствительность немедленного типа. Обусловлена освобождением активных субстанций (в основном гистамина) из тучных клеток, сенсибилизрованных IgE-антителами, при связывании ими аллергена. Примеры - сенная лихорадка, пищевые аллергии. ● 2-й тип — гиперчувствительность, обусловленная цитотоксическим эффектом антител (IgG), вовлекающих комплемент или эффекторные клетки. ○ Аллоимммунные гемолитические заболевания ○ Гемолитические процессы, связанные с лекарственной гиперчувствительностью ● 3-й тип — иммунокомплексная реакция. Обусловлена провоспалительным действием растворимых иммунных комплексов, которые переходят в нерастворимое состояние (феномен Артюса-Сахарова). ● 4-й тип — гиперчувствительность замедленного типа. Связана с активностью провоспалительных Т-лимфоцитов и активируемых ими макрофагов, а также цитокинов, секретируемых названными клетками (реакция Манту).
55. Роль тучных клеток, Th2-лимфоцитов и продуцируемых ими цитокинов в реакциях гиперчувствительности. Тучные клетки: При гиперчувствительности анафилактического типа при первичном контакте с антигеном образуются IgE прикрепляющиеся Fc-фрагментом к базофилам и тучным клеткам. Повторное введение антигена вызывает его связывание с антителами и дегрануляцию клеток с выбросом медиаторов воспаления, прежде всего гистамина - вики. ● формируют фон для развития Th2-иммунного ответа ● дегрануляция (гистамин, гепарин итд) ● выброс эйкозаноидов - расширение сосудов, спазм гладкой мускулатуры ● выброс цитокинов - привлечение эозинофилов, базофилов, нейтрофилов Th2: ● выброс цитокинов - привлечение эозинофилов, базофилов, нейтрофилов ● развитие тучных клеток ● переключение на синтез IgE
56. Роль IgE в развитии гиперчувствительности I типа ● IgE-антитела обладают способностью фиксироваться на тучных клетках или базофилах. При присоединении аллергенов с реагинами на поверхности тучных клеток или базофилов, из них выделяются физиологически активные субстанции (гистамин). Продукты тучных клеток и базофилов, действуя на клетки-мишени, вызывают ответ, неадекватный по интенсивности и по “целевой” направленности исходному воздействию. Этот ответ обозначается как аллергическая реакция
57. Какое событие на клеточном уровне является причиной аллергических реакций I типа? ● Выброс активных молекул тучными клетками, сенсибилизированными IgE-антителами при связывании ими аллергенов.
58. Какие клетки служат основой защиты от заражения макропаразитами? ● эозинофилы (в кровотоке), тучные клетки (в тканях). (10 лекция Д.В.Купраша, 01:14:52). ● 59. Физиологическая функция эозинофилов и механизмы киллинга? ( Ярилин 2010, стр 57) ● защита от макропаразитов: прикрепляются к поверхности паразита и дегранулируют белки ответственные за поражение: например, основный белок (МВР) и эозинофильный катионный белок (ЕСР). Эти молекулы встраиваются в мембрану паразитов и разрушают ее. ● защита от вирусов: катионный белок (ЕСР) и нейротоксин (EDN) - рибонуклеазы. - голубым отмечено самое важное ● Функция эозинофилов в норме заключается в регуляции развития тучных клеток и морфогенетических процессов, связанных с беременностью и половым циклом у самок. ● положительная селекция Т-клеток в тимусе. ● запуск Th2-зависимых иммунных процессов, в частности аллергических. ● Поддержание клеток памяти в их нишах.
60. Особенности альтернативного, лектинового и классического путей комплемента. (лекция 4 слайд 9) 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|