
|
|
Дополнительные сведения и гипотезы: ТератокарциномаПоловая клетка млекопитающих или ранний бластомер содержит в себе всю информацию, необходимую для последующего развития. Что может случиться, если такая клетка станет злокачественной? В этом случае возникает тератокарцинома. Опухоли этого типа иногда спонтанно развиваются в яичнике или в семеннике. Образование тератокарцином можно вызвать экспериментально, имплантируя бластоцисту не в матку, а в иное место (например, внутрь соединительнотканной капсулы, окружающей почку). Независимо от того, возникла ли тератокарцинома спонтанно или в результате экспериментального вмешательства, она содержит популяцию недифференцированных стволовых клеток, которые по биохимическим свойствам и по способностям к развитию похожи на клетки внутренней клеточной массы (Graham, 1977). Эти стволовые клетки (называемые клетками эмбриональной карциномы) не только делятся, но могут также дифференцироваться в самые разнообразные ткани, в том числе в кишечный и дыхательный эпителий, мышечную, нервную, хрящевую и костную ткани (рис. 6.44). Образующиеся дифференцированные клетки больше не делятся и потому не являются злокачественными. Итак, эти опухоли могут дать начало самым разнообразным тканям, встречающимся в организме. Эмбриональные карциномы можно переносить из одной мыши в другую; их клетки можно индуцировать к образованию эмбриоидных тел. инъецируя эти клетки в перитонеальную (брюшную) полость.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 222 ___________ ГЛАВА 6___________________
где затем появляются аморфные скопления дифференцированных клеток (в том числе кроветворные колонии) в «энтодерме» (рис. 6.45). Было показано (Adamson et al., 1977), что эта «энтодерма» способна синтезировать α-фетопротеин, белок, обычно синтезируемый нормальными энтодермальными клетками млекопитающих. Следовательно, клетки эмбриональной карциномы имитируют раннее развитие млекопитающих, но развитие формируемой ими опухоли имеет случайный, неупорядоченный характер. Эти опухоли, вероятно, происходят из половых клеток, однако по своим биохимическим, морфологическим и поверхностным свойствам образующие ее клетки напоминают бластомеры внутренней клеточной массы. Стюарт и Минц (Stewart, Mintz, 1981) получили мышь из клеток, часть которых происходила из стволовой клетки тератокарциномы! Стволовые клетки из тератокарциномы мышей линии агути культивировали в течение нескольких клеточных поколений и убедились, что они сохраняют хромосомный набор, характерный для родительской мыши. Затем изолированные стволовые клетки этого типа инъецировали в бластоцисты белых мышей. Бластоцисты переносили в матку приемной матери, которая родила живых мышат. Шкурки у некоторых мышат были двухцветными; это свидетельствовало о включении опухолевой клетки в развитие зародыша (рис. 6.46). Более того, когда химерную мышь спаривали с мышью, несущей соответствующий маркер, некоторые из рожденных ею мышат имели черты, характерные для фенотипа опухолевого «родителя». Злокачественная клетка эмбриональной карциномы образовала многие, если не все, типы нормальных соматических клеток и даже нормально функционирующие половые клетки! Если спаривали
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
Рис. 6.46. Введение клетки эмбриональной карциномы в бластоцисту мыши. А. Микропипетка со стволовой клеткой тератокарциномы приближается к бластоцисте, прочно удерживаемой обычной пипеткой. Б. Инъекция стволовой клетки во внутреннюю клеточную массу. В. Мыши, развившиеся в опыте, в котором стволовая клетка тератокарциномы из черной линии мышей интегрировалась во внутреннюю клеточную массу бластоцисты мышей белой линии. На голове и спине мышей можно видеть черный мех, указывающий на то, что по крайней мере пигментные клетки были опухолевого происхождения. (А и Б — с с любезного разрешения K. Illmense; В – из Papaioannou, 1979; фотография с любезного разрешения V.E. Papaioannou. )
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 224 ГЛАВА 6
мышей, получивших опухолевую клетку от одного из родителей, то в потомстве оказывались мыши, гомозиготные по большому числу генов опухолевой клетки (рис. 6.47). Поскольку культивирование ранних зародышей млекопитающих является трудной задачей, клетки эмбриональной карциномы дают исследователям уникальную возможность для анализа раннего развития млекопитающих. Кроме того, благодаря способности клеток эмбриональной карциномы мутировать в культуре при инъекции таких клеток в бластоцисты можно также создать мышей, мутантных почти по любому гену. На этом мы завершим обзор основных черт развития животных и обратимся к механизмам, которые делают возможным это развитие. Во втором томе книги мы рассмотрим различные способы, с помощью которых осуществляется дифференцировка клеток, а третий том будет посвящен вопросам взаимодействия клеток между собой при формировании тканей и органов.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
________________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ МЕЗОДЕРМА И ЭНТОДЕРМА____________________________________225
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 226_______________ ГЛАВА 6_______________________________________________________________________________
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
Оглавление
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 228_______________ ОГЛАВЛЕНИЕ___________________________________________________________________________

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|



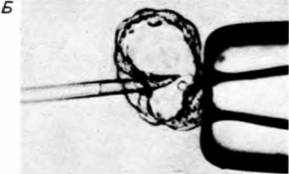

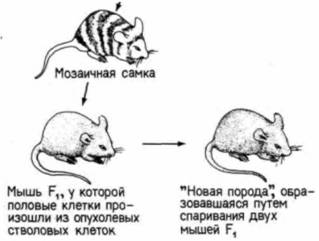 Рис. 6.47. Схема опыта с получением мышей, гены которых принадлежат главным образом опухолевым клеткам. Из злокачественной карциномы мыши изолировали линию стволовых клеток и вводили их в бластоцисты мыши другой линии. Химерные бластоцисты помещали для развития в матку приемной матери. Если опухолевые клетки интегрируются в бластоцисту, то многие из клеток у родившейся мыши происходят из клеток опухоли. Если из стволовых клеток опухоли произошли и половые клетки, то мозаичную мышь можно спарить с нормальной мышью и получить потомство F1. Мыши F1 должны быть гетерозиготными по всем хромосомам опухолевой клетки. Спаривание между мышами F1 дает мышей F2, имеющих гомозиготные гены, происходящие из опухолевых клеток. (По Stewart, Mintz, 1981.)
Рис. 6.47. Схема опыта с получением мышей, гены которых принадлежат главным образом опухолевым клеткам. Из злокачественной карциномы мыши изолировали линию стволовых клеток и вводили их в бластоцисты мыши другой линии. Химерные бластоцисты помещали для развития в матку приемной матери. Если опухолевые клетки интегрируются в бластоцисту, то многие из клеток у родившейся мыши происходят из клеток опухоли. Если из стволовых клеток опухоли произошли и половые клетки, то мозаичную мышь можно спарить с нормальной мышью и получить потомство F1. Мыши F1 должны быть гетерозиготными по всем хромосомам опухолевой клетки. Спаривание между мышами F1 дает мышей F2, имеющих гомозиготные гены, происходящие из опухолевых клеток. (По Stewart, Mintz, 1981.)

