|
|
Дополнительные сведения и гипотезы: Эволюция развития головного мозга
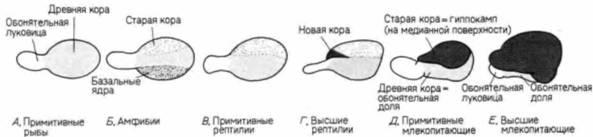
Эволюция коры головного мозга — один из наиболее интересных разделов анатомии позвоночных. Доли мозга возникают как парные выросты переднего мозга. У рыб, например, они первоначально функционируют как органы обоняния. У амфибий и рептилий древнюю кору (палеопаллиум) головного мозга покрывают старая кора (архипаллиум) и полосатое тело (corpus striatum) (рис. 5.18). Архипаллиум. по-видимому, связан с «эмоциональными» типами поведения, тогда как палеопаллиум и полосатое тело имеют отношение к автоматическим «инстинктивным» реакциям. У птиц полосатое тело развито очень хорошо. У более высокоорганизованных рептилий появляются первые нейроны, связанные с новой корой (неопаллиумом, или неокортексом). Эволюция мозга млекопитающих, и особенно человека, характеризуется необычайно интенсивным ростом и распространением новой коры, оттесняющей внутрь архипаллиум, который формирует гиппокамп (ответственный за сексуальное и агрессивное поведение). Мозолистое тело функционирует как реле для переключения некоторых непроизвольных реакций. Неокортекс становится центром обучения, памяти и интеллекта. У птиц нет новой коры, и у видов, способных к обучению, его функцию выполняет гиперстриатум (Romer, 1976). Развитие новой коры у человека занимает чрезвычайно большой период времени. Фактически у ребенка после его рождения мозг продолжает развиваться с такой же скоростью, как у плода (Holt et al., 1975). Портманн (Porlmann. 1941. 1945) полагает, что по сравнению с другими приматами у человека очень короткий период беременности. Сравнивая развитие плода у человека и других приматов, он определил, что беременность у женщины должна была бы продолжаться 21 мес. Портманн и Гоулд (Gould. 1977) считали, что именно такова и есть продолжительность беременности у человека, поскольку все мы в течение первого года жизни являемся в сущности внематочными плодами. Это утверждение основывается на том, что и после рождения мозг ребенка продолжает расти чудовищно быстрым темпом, характерным для плода. Ни одна женщина не могла бы родить 21-месячного младенца, потому что его голова не могла бы пройти через родовые пути. Роды должны состояться в то время, когда голова у младенца еще достаточно мала, чтобы пройти через материнский таз. Поэтому человеческие детеныши рождаются в конце девятого, а не двадцать первого месяца беременности. Кроме того, именно в течение этого времени мы впервые подвергаемся влиянию окружающего мира. Наша развивающаяся нервная система получает существенный выигрыш от воздействия на нее столь сильной стимуляции в течение первого года жизни ребенка.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 160 ГЛАВА 5 типы нейронов. В головном мозге человека содержится более 100 миллиардов нервных клеток (нейронов), связанных с более чем триллионом вспомогательных глиальных клеток. Клетки, остающиеся неотъемлемыми компонентами выстилки нервной трубки, становятся эпендимными клетками. Из них могут образоваться как предшественники нейронов, так и глиальные клетки (рис. 5.19). Полагают, что дифференцировка этих клеток-предшественников в значительной степени детерминируется окружением, в которое они попадают (Rakic, Goldman, 1982), и что по крайней мере в некоторых случаях данная клетка-предшественник может формировать как нейроны, так и глиальные клетки (Turner, Cepko, 1987). Типы нейронов и глиальных клеток чрезвычайно разнообразны (это можно видеть из сравнения относительно мелкой клетки зернистого слоя и огромного нейрона Пуркинье). У некоторых нейронов образуется лишь небольшое число цитоплазматических участков, в которых другие клетки могут передавать им электрические импульсы, тогда как другие нейроны формируют обширные области, доступные для этой цели. Тонкие отростки нервных клеток, служащие для восприятия электрических импульсов, называются дендритами(рис. 5.20). Нейроны коры головного мозга новорожденного имеют очень мало дендритов, и одним из самых важных событий первого года его жизни является увеличение числа таких рецепторных участков в этих нейронах. В течение первого года жизни ребенка у каждого нейрона в коре головного мозга образуется поверхность дендритов, достаточная для размещения на ней 100 000 связей с другими нейронами. В среднем один нейрон коры головного мозга соединяется с 10 000 других нервных клеток. Этот характер нервных связей делает возможным функционирование коры головного мозга как центра обучения, мышления и памяти, развития способностей к обобщениям и к произвольным ответам на воспринимаемые и расшифрованные ею стимулы. Другой важной структурой развивающегося нейрона является аксон. Если дендриты часто очень многочисленны и не распространяются далеко от тела нервной клетки (или сомы), то аксоны простираются на несколько футов ( 1 фут = 30.5 см). Так, частоту сердечных сокращений регулируют нервы, клеточные тела которых находятся в продолговатом мозге. Болевые рецепторы большого пальца ноги воспринимают импульсы, которые должны пройти долгий путь до спинного мозга. Одна из фундаментальных концепций нейробиологии заключается в том, что аксон представляет собой непрерывное продолжение тела нервной клетки. Еще в конце прошлого столетия существовали многочисленные конкурирующие теории образования аксона. Шванн, один из основоположников клеточной теории, считал, что многочисленные нервные клетки соединяются между собой в цепочку, формируя аксон. Гензен, открывший зародышевый узелок (названный впоследствии его именем), считал, что аксон формируется вокруг предсуществующих цитоплазматических тяжей между клетками. Вильгельм Гис (His, 1886) и Сантьяго Рамон-и-Кахал (Ramon у Cajal, 1890) утверждали, что аксон, безусловно, представляет собой вырост нервной сомы (хотя и чрезвычайно длинный). Позже Гаррисон (Harrison. 1907) продемонстрировал справедливость теории выроста в изящном эксперименте, которым одновременно были заложены основы нейробиологии развития и метода культуры тканей. Гаррисон изолировал участок нервной трубки из зародыша лягушки длиной в три миллиметра. На этой стадии (вскоре после слияния нервных валиков) еще нет видимой дифференцировки аксонов. Он поместил нейробласты на покровное стекло в каплю лимфы лягушки и перевернул стекло над углублением в предметном стекле так, чтобы было возможно наблюдать за тем, что происходит в этой «висячей капле». И Гаррисон увидел (рис. 5.21), что аксоны образуются как выросты нейробластов, удлиняющиеся со скоростью примерно 56 мкм/ч. Движение выроста осуществляется с помощью ведущего кончика аксона, называемого конусом роста (рис. 5 22). Конус роста движется не по прямой. он как бы нащупывает свой путь по субстрату. Движение конуса осуществляется посредством удлинения и сокращения заостренных филоподий, называемых микрошипами. Эти филоподии содержат микрофиламенты, ориентированные параллельно длинной оси аксона. (Микрофиламенты обусловливают процессы миграции клеток некоторых типов и всегда принимают участие в сократительной активности клеток.) Обработка нейронов цитохалазином В подавляет их дальнейшее продвижение (Yamada et al.. 1971). В пределах самого аксона прочность его структуры обеспечивается микротрубочками: если нейробласт поместить в раствор колхицина, то аксон будет втягиваться в тело клетки (рис. 5.23). Таким образом, развивающийся нейрон сохраняет черты, уже обсуждавшиеся нами при изучении формирования нервной трубки, а именно удлинение клетки при участии микротрубочек и изменение формы ее апикального конца с помощью микрофиламентов. В наиболее активно мигрирующих клетках «исследующие» филоподии прикрепляются к субстрату и тянут за собой остальную часть клетки. Учитывая длину аксона, можно понять, что ему такую операцию проделать было бы трудно. Поэтому сократительный аппарат в филоподиях аксона является, по-видимому, только исследующим, а уд-
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. _______________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ ЭКТОДЕРМА 161
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. ________________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ/ ЭКТОДЕРМА________________________________________ 163
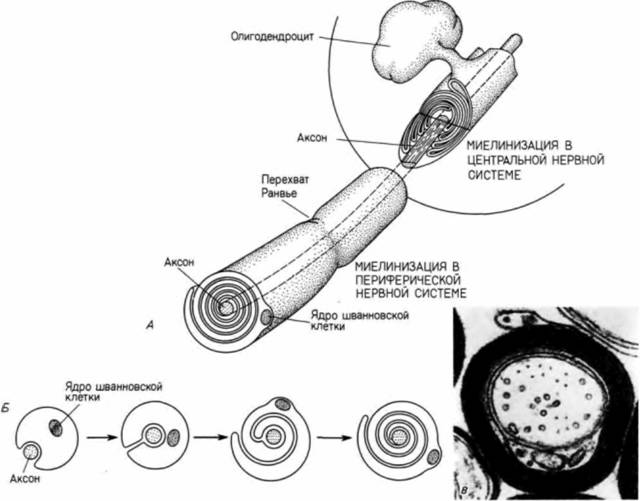
линение аксона осуществляется при помощи перетекающей по нему цитоплазмы. Нейроны передают электрические импульсы от одного участка к другому. Эти импульсы обычно проходят через дендриты в тело нейрона, где они фокусируются в аксоне. Чтобы предотвратить рассеяние электрического сигнала и облегчить его проведение, аксон в центральной нервной системе через определенные промежутки на всем его протяжении изолируется от других аксонов в нерве отростками глиальных клеток, называемых олигодендроцитами.Олигодендроцит как бы обвертывает собой развивающийся аксон. Затем он продуцирует специализированную клеточную оболочку, богатую основным белком миелином, которая окружает в виде спирали лежащий в ее центре аксон (рис. 5.24). Специализированная мембрана называется миелиновой оболочкой.(В периферической нервной системе миелинизацию аксонов осуществляют шванновские глиальные клетки.) Миелиновая оболочка имеет существенное значение для правильного функционирования нейрона: демиелинизация нервных волокон связана с некоторыми серьезными нарушениями умственной деятельности; иногда процесс демиелинизации приводит к летальному исходу. Аксон должен быть также специализирован для секреции нейромедиаторов, способных проходить через небольшие пространства (синаптические щели), отделяющие аксон нервной клетки от поверхности его клетки-мишени (ее тела, дендритов, аксона, воспринимающего нейрона или рецепторов какого-либо периферического органа). Таким образом, развивающийся нейрон должен продуцировать специфические ферменты для синтеза и разрушения нейромедиаторов. Развитие нейрона означает не только структурную, но и молекулярную дифференцировку. Развитие глаза ДИНАМИКА РАЗВИТИЯ ГЛАЗА. Индивидуум приобретает знания об окружающем его мире с помощью органов чувств. В этом разделе мы сосредоточим внимание на развитии глаза, потому что для него, вероятно, более, чем для какою-либо другого органа тела, необходимы точность и совершенство координации развития всех его компонентов. Развитие органа зрения начинается в стенке будущего промежуточного мозга. У человека это происходит на 22-й день развития, когда в стенке нервной трубки образуются латеральные выпячивания, которые впоследствии станут промежуточным мозгом. Дифференциальный рост выпячиваний приводит к образованию глазных пузырей,связанных с промежуточным мозгом глазными стебельками.Позже глазные пузыри приходят в контакт с эктодермальным покровным эпителием зародыша и индуцируют образование в нем хрусталиковых плакод(рис.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 164 ГЛАВА 5
5.25) 1. Эта индукция является специфической, поскольку изолированные глазные пузыри, имплантированные под эпителий любой области головы, будут вызывать в нем образование хрусталиковой ткани вместо эпидермальных клеток. В отсутствие контакта с глазным пузырем хрусталик не образуется. Необходимость тесного контакта между глазными пузырями и покровным эпителием подтверждается не только в экспериментах с трансплантацией глазных пузырей, но и нарушениями в развитии глаз у некоторых мутантов. Например, у мутантов мыши eyeless глазные пузыри не соприкасаются с покровным эпителием и формирование глаза прекращается (Webster et al., 1984). Сформировавшись, хрусталиковая плакода в свою очередь вызывает изменения в глазном пузыре, передняя стенка которого инвагинирует и пузырь превращается в двустенную глазную чашу (рис. 5.26). По мере продолжения инвагинации глазной стебелек, соединяющий глазную чашу и головной мозг, сужается и связь между ними редуцируется до узкой щели. Одновременно с этим два слоя глазной чаши начинают дифференцироваться в разных направлениях. Клетки наружного слоя продуцируют пигмент и в конце концов образуют пигментированный слой, называемый пигментной сетчаткой. Число клеток внутреннего слоя быстро увеличивается, и эти клетки дают начало множеству светочувствительных (фоторецепторных) нейронов, глиальных клеток, промежуточных нейронов и ганглиозных клеток. Все эти клетки в совокупности составляют нейральную ретину,или сетчатку. Аксоны ганглиозных клеток нейральной сетчатки встречаются у основания глаза и далее проходят по глазному стебельку, который после этого начинают называть зрительным нервом.
1 Процессы индукции, участвующие в формировании глаза, подробно рассматриваются в гл. 8 и 16.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. ________________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ. ЭКТОДЕРМА________________________________________ 165
ДИФФЕРЕНЦИРОВКА НЕЙРАЛЬНОЙ СЕТЧАТКИ. Подобно тому как это происходит в коре головного мозга и мозжечка, в сетчатке по мере ее развития разные нейроны группируются в слои свето- и цветочувствительных фоторецепторных клеток, тел ганглиозных клеток и биполярных промежуточных нейронов, передающих электрические стимулы от палочек и колбочек к ганглиозным клеткам (рис. 5.27). В дополнение к ним имеются многочисленные глиальные клетки, поддерживающие целостность сетчатки, а также амакриновые и горизонтальные нейроны, которые передают электрические импульсы в горизонтальном направлении. На ранних стадиях развития сетчатки образование ее слоев обусловлено делением клеток герминативного слоя, их миграцией и дифференциальной гибелью. Формирование этой сложно структурированной ткани представляет собой одну из наиболее интенсивно изучаемых проблем нейробиологии развития. Недавно было показано (Turner, Cepko, 1987), что любая единичная клетка-предшественник нейробласта может дать начало по меньшей мере трем типам нейронов или двум типам нейронов и одному типу глиальных клеток. Этот анализ был осуществлен с помощью очень хитроумного способа мечения клеток, происходящих из одной конкретной клетки-предшественника. Новорожденным крысятам (с еще развивающейся сетчаткой) в заднюю стенку глаза инъецировали вирус, который мог включаться в ДНК клеток глаза. Этот вирус содержал ген β-галактозидазы (отсутствующий в сетчатке глаза крысы), который должен был экспрессироваться в инфицированных клетках. Через месяц после введе-
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 166 ГЛАВА 6
ния вируса сетчатку удаляли и окрашивали красителем, выявляющим присутствие β-галактозидазы. В синий цвет должны были окраситься только потомки инфицированных клеток. На рис. 5.28 показана одна из полос клеток, происшедших от инфицированной клетки-предшественника. Краситель виден в пяти палочках, биполярном нейроне и мюллеровской глиальной клетке. Из трех основных типов нейронов сетчатки (ганглиозные, биполярные и фоторецепторные) фоторецепторные палочки и колбочки, вероятно, завершают свою дифференцировку последними. По мере развития этих наружных нейронов тело каждого из них образует вырост, содержащий некоторые специализированные органеллы. Эти органеллы удлиняют вырост и определяют размер и форму фотореактивных областей (Detwiler, 1932). Клеточные мембраны наружных нейронов складываются, образуя мешки, в которых размещаются фоторецептивные (зрительные) пигменты. Свет индуцирует химические изменения этих пигментов, приводящие в конечном счете к изменению мембранного потенциала. Изменение мембранного потенциала влияет на высвобождение нейромедиаторов из группы биполярных нейронов, которые переключают электрический сигнал на ганглиозные клетки. Аксоны этих клеток, соединяясь в пучки, образуют зрительный нерв, по которому полученная информация передается в головной мозг (Fesenko et al., 1985; Stryer, 1986). 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|