|
|
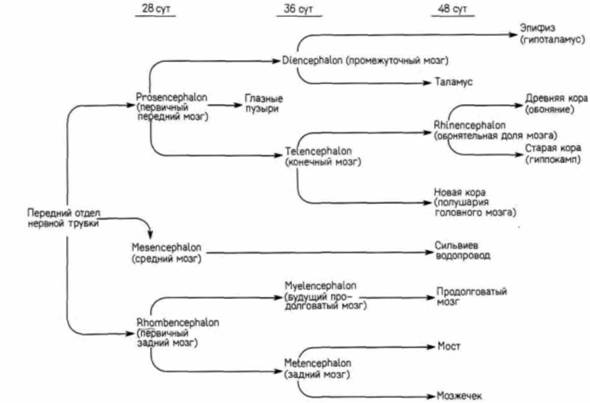
Дифференцировка нервной трубкиФОРМИРОВАНИЕ ОТДЕЛОВ ГОЛОВНОГО МОЗГА. Разные участки нервной трубки дифференцируются в разные отделы центральной нервной системы одновременно тремя различными путями. На анатомическом уровне в нервной трубке возникают расширения и сужения, а ее полости образуют камеры головного и спинного мозга. На гистологическом (тканевом) уровне клеточные популяции в стенке нервной трубки перераспределяются разными способами, формируя разные функциональные отделы головного и спинного мозга. И наконец, на цитологическом (клеточном) уровне сами клетки нейрального эпителия дифференцируются с образованием многочисленных типов нейронов и опорных (глиальных) клеток. Ранние стадии развития головного мозга у большинства позвоночных очень сходны, но, поскольку головной мозг человека представляется наиболее интересным органом в животном царстве, мы сосредоточим внимание на развитии того, что, как предполагают, делает человека — Нотo —человеком разумным Homo sapiens. У раннего зародыша млекопитающих нервная трубка на всем ее протяжении прямая. Однако даже раньше, чем сформируется задний участок трубки, самая передняя ее часть претерпевает чрезвычайно резкие изменения. Она раздувается, подразделяясь на три первичных мозговых пузыря (рис. 5.10): первичный передний мозг (prosencephalon), средний мозг(mesencephalon) и первичный задний мозг (rhombencephalon). К тому времени, когда замкнется задний конец нервной трубки, по бокам развивающегося переднего мозга образуются вторичные расширения — глазные пузыри. Кроме того, будущий головной мозг изгибается таким образом, что места изгиба отмечают границы полостей мозга. Два основных изгиба называются головным и затылочным (шейным). Первичный передний мозг затем подразделяется на передний конечный мозг (telencephalon) и расположенный каудальнее его промежуточный мозг (diencephalon) (рис. 5.11). Конечный мозг впоследствии формирует большие полушария головного мозга, промежуточный — таламическую и гипоталамическую области, а также область, получающую нервные импульсы от глаз; средний мозг на отделы не подразделяется, а его полость в конце концов становится третьим желудочком, или сильвиевым водопроводом. Первичный задний мозг подразделяется на будущий продолговатый мозг (myelencephalon) и лежащий впереди него собственно задний мозг (metencephalon). Myelencephalon в конечном счете становится продолговатым мозгом (medulla ohlongata),
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 154_______________ ГЛАВА 5__________
нейроны которого формируют нервы, регулирующие деятельность дыхательной, желудочно-кишечной и сердечно-сосудистой систем. А задний мозг формирует мозжечок (cerebellum) — отдел мозга, ответственный за координацию движений, поддержание позы и чувство равновесия. Развитие специализированных отделов головного мозга суммировано на рис. 5.12. Процесс образования мозговых пузырей у раннего зародыша характеризуется чрезвычайно высокой скоростью и, кроме того, тем, что этиогромные вздутия образуются в основном за счет увеличения размеров полостей, а не за счет утолщения их стенок. У куриного зародыша объем головного мозга между третьими и пятыми сутками развития увеличивается в 30 раз. Полагают, что такое быстрое увеличение
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. ________________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ. ЭКТОДЕРМА_______________________________________ 155
размеров вызвано давлением на стенки нервной трубки заключенной в ней жидкостью. Давление этой жидкости по логике вещей должно было бы уменьшаться в результате ее проникновения в полость спинного мозга, однако этого не происходит. Несколькими авторами (Schoenwolf, Desmond, 1984; Desmond, Shoenwolf, 1986) было показано, что в нервной трубке куриного зародыша до замыкания заднего нейропора возникает сужение, отделяющее будущий головной мозг от спинного (рис. 5.13). (Такая же преграда образуется и у зародыша человека: Desmond, 1982.) Если у куриного зародыша снять давление жидкости в передней части такой перегороженной нервной трубки, то скорость увеличения объема головного мозга значительно снижается и он будет содержать гораздо меньше клеток, чем головной мозг интактных контрольных зародышей. В норме после первоначального быстрого увеличения объемов желудочков (полостей) головного мозга перегороженный участок нервной трубки снова открывается. АРХИТЕКТОНИКА ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ НА ТКАНЕВОМ УРОВНЕ. Исходно нервная трубка образована слоем герминативного нейрального эпителия толщиной в одну клетку. Этот слой представляет собой быстро делящуюся клеточную популяцию. Сауэр (Sauer. I935) и другие исследователи показали, что клетки этой популяции располагаются в один слой; каждая клетка простирается от края, обращенного к просвету нервной трубки, до ее наружного края. Однако ядра клеток находятся на разных уровнях, и потому создается впечатление, что стенка нервной трубки многослойна. Такое положение ядер зависит от стадии клеточного цикла (рис. 5.14). Синтез ДНК (S-фаза) происходит тогда, когда ядро клетки располагается у ее наружного конца: по мере приближения к фазе митоза оно мигрирует по направлению к просвету трубки. Митоз осуществляется вблизи того конца клетки, который обращен к просвету нервной трубки. В период раннего развития 100% клеток нервной трубки включают радиоактивный тимидин в ДНК (Fujita, 1964). Однако вскоре некоторые клетки перестают включать предшественники ДНК, показывая тем самым, что они более не участвуют в синтезе ДНК и в митозе. Эти клетки представляют собой молодые нейроны или глиальные (опорные) клетки; они мигрируют к периферии нервной трубки, где происходит их дальнейшая дифференцировка (Fujita. 1966; Jacobson. 1968). Она зависит от положения, которые займут эти нейробласты, оказавшись вне области делящихся клеток (Jacobson, 1978; Letourneau, 1977). В то время как клетки, лежащие у просвета нервной трубки, продолжают делиться, мигрирующие клетки образуют второй слой вокруг исходного слоя клеток нервной трубки. Этот второй слой постепенно утолщается по мере того, как все большее число клеток добавляется к нему из герминативною нейрального эпителия; этот слой называют плащевым(рис. 5.15. А), агерминативный эпителий теперь но-
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 156______________ ГЛАВА 5_______________________________________________________________________________
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ. ЭКТОДЕРМА 157 сит название эпендимного слоя.Дифференцировка клеток плащевого слоя приводит к образованию как нейронов, так и глиальных клеток. Нейроны устанавливают между собой связи и посылают в направлении от просвета аксоны, образуя вблизи него бедный клетками краевой слой. В конце концов глиальные клетки заключают аксоны этого слоя в миелиновые футляры, придающие им беловатую окраску. Поэтому плащевой слой, содержащий тела нейронов, часто называют серым веществом, асодержащий их аксоны краевой слой белым веществом В спинном и продолговатом мозге основная трехкомпонентная структура — эпендимный, плащевой и краевой слои — сохраняется в течение всего развития. Серое вещество (плащ, или мантия) постепенно принимает форму бабочки, окруженной белым веществом, и оба они заключены в соединительнотканный чехол. По мере дифференцировки нервной трубки в ней образуется продольная борозда(sulcus limitans), делящая ее на дорсальную и вентральную половины. Дорсальная половина получает информацию от чувствительных (сенсорных) нейронов, вентральная участвует в осуществлении различных двигательных (моторных) функций. В головном мозге, однако, миграция клеток, дифференциальный рост и селективная гибель клеток вызывают модификацию трехзонной структуры, особенно в мозжечке (рис. 5.15, Б—Г). В глубоких слоях мозжечка вблизи просвета из нейробластов формируются скопления нейронов, называемые ядрами. Каждое ядро работает как функциональная единица, служащая промежуточной ретрансляционной станцией между наружными частями мозжечка и другими частями мозга. Другие нейробласты серого вещества мигрируют по наружной поверхности развивающегося мозжечка, образуя новый герминативный слой вблизи наружной границы нервной трубки. Нейробласты, происходящие из клеток этого слоя, мигрируют обратно в развивающееся белое вещество мозжечка, в котором формируют зернистый слой.Из исходного эпендимного слоя возникают разнообразные нейроны и глиальные клетки. включая отличающиеся от других нейронов крупные клетки Пуркинье.Каждый нейрон Пуркинье имеет огромный дендритный аппарат,который подобен опахалу над луковицеобразным телом клетки (рис. 5.16. А). Типичная клетка Пуркинье может образовывать до 100 000 синапсов с другими нейронами — больше, чем любой другой из изученных нейронов. У каждого нейрона Пуркинье имеется тонкий аксон,соединяющий его с другими клетками в глубоких ядрах мозжечка. Таким образом, развитие пространственной организации мозжечка является решающим условием его правильного функционирования. В сущности все импульсы регулируют активность клеток Пуркинье. которые представляют собой единственные эфферентные нейроны коры мозжечка. Для осуществления такой регуляции необходимо, чтобы дифференцировка соответствующих клеток происходила в надлежащем месте и была координированной во времени. Пониманию механизмов, обусловливающих упорядоченность расположения нейронов мозжечка в пространстве, способствовал анализневрологических мутаций у мышей. Известно свыше 30 мутаций, влияющих на распределение этих нейронов. Мутация staggerer, по-видимому, вызвана дефектами клеток Пуркинье, которые умутантов мельче, чем у нормальных мышей, и имеют короткие слабоветвящиеся дендриты (рис. 5.16. Б). Клетки зернистого слоя у мутантов образуются своевременно и мигрируют нормально, но затем погибают. По мнению некоторых авторов (Sidman, 1974: Sotelo, Changcaux, 1974), эти клетки погибают потому, что им не удается установить связь с дендритами клеток Пуркинье. Мыши с дефектами клеток Пуркинье меньше по размерам, для них характерна своеобразная шатающаяся походка, и они непрерывно дрожат. Многие мутации, обусловливающие дефекты клеток мозжечка, были обнаружены по легко распознаваемому фенотипу таких мутантов. У них всегда бывает нарушено чувство равновесия и затруднены движения. Фенотип мутантов weaver (качание) схо-
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 1 58 ГЛАВА 5
ден с фенотипическим проявлением мутации staggerer; строение клеток Пуркинье у них также аномально. Однако у мышей weaver остановка роста клеток Пуркинье представляет собой вторичное явление — она обусловлена, по-видимому, генетическим дефектом клеток зернистого слоя. Это было доказано в опытах «конструирования» химерных мышей из зародышей дикого типа и мутантов weaver (Goldowitz, Mullen, 1982). У химерных мышей клетки дикого типа можно обнаружить в мозжечке по низкому содержанию в них ß-глюкуронидазы (выявляемой гистохимическим методом) и по иной форме ядер. У мутантов меньше клеток Пуркинье, чем у мышей дикого типа, и локализация этих клеток в коре мозжечка аномальна. У химерных зародышей некоторые из нейронов Пуркинье также располагались не там, где они должны были находиться. Эти эктопические нейроны представляли собой клетки как того, так и другого генотипа. Однако все клетки зернистого слоя у мышей дикого типа находились там, где им положено, тогда как у мутантных зародышей эти клетки были случайно распределены по коре мозжечка (рис. 5.17. А). Таким образом, дефект у мышей weaver заключается, по-видимому, в отсутствии у клеток зернистого слоя способности мигрировать из наружного герминативного слоя в ту область коры мозжечка, где они должны находиться. В зернистый слой нейробласты обычно мигрируют, двигаясь по длинным отросткам глиальных клеток Бергмана (Rakic, Sidman, 1973). Хаттеном и его коллегами (Hatten et al.. 1986) было показано, что у мутантов weaver нейробласты неспособны узнавать клетки Бергмана как субстрат, по которому они должны мигрировать. В опытах этих исследователей в разных сочетаниях соединяли клетки зернистого слоя и клетки глии нормальных и мутантных зародышей (рис. 5.17, Б). Клетки от мышей дикого типа прочно слипались с глией зародышей обоих генотипов (как это обычно и бывает, когда клетки мигрируют). Однако у мутантных зародышей они не приближались ни к глиальным клеткам зародышей дикого типа, ни к глиальным клеткам зародышей weaver. Эти данные убедительно подтверждают предположение, что генетический эффект мутации weaver заключается в неспособности будущих клеток зернистого слоя узнавать клетки Бергмана и мигрировать по их поверхности. Трехзонная структура мозга в полушариях также претерпевает существенные изменения. Некоторые нейробласты из плащевого слоя мигрируют через белое вещество и образуют вторую зону нейронов, называемую новой корой (неопаллиумом, или неокортексом). Эта новая кора мозга затем подразделяется на шесть слоев, содержащих тела нейронов. Дифференцировка этих слоев продолжается после рождения ребенка в течение первой половины детства.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ. ЭКТОДЕРМА 159 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|