
|
|
Ближний и дальний транспорт водыПодавляющая часть мощного водного тока, непрерывно проходящего через растение, непосредственно в биохимических превращениях не участвует. В фотосинтезе, дыхании, гидролитических и других реакциях используется не более 1 % поглощенной воды. Активное проявление жизнедеятельности возможно только в водной среде, которая должна постоянно циркулировать по организму. За счет циркуляции вода доставляет клеткам необходимые для их функционирования вещества и одновременно удаляет продукты жизнедеятельности, в том числе токсические отбросы. Именно благодаря такой циркуляции осуществляется взаимосвязь между отдельными частями клеток, между отдельными клетками в пределах ткани, между различными тканями в пределах органа, наконец, между различными органами и тем самым обеспечиваются взаимосвязь физиологических процессов, поддержание гомеостаза и функционирование организма как единого целого. Транспорт воды является важнейшей физиологической функцией, в регулировании которой самое непосредственное участие принимают не только внешние, но и главным образом внутренние факторы. Концевые двигатели восходящего тока – нижний (корневое давление) и верхний (присасывающее действие транспирации) взаимосвязаны, а также имеют связь с другими процессами жизнедеятельности, что обеспечивается сложной системой эндогенной регуляции. Транспорт воды в тканях одного органа называют ближним, а между органами – дальним. Ближний транспорт идет по неспециализированным тканям, для дальнего – имеются специальные ткани – проводящие. Основная масса воды двигается вверх по растению: из корней в листья (восходящий ток); небольшая часть – вниз (нисходящий ток). Восходящий ток идет по сосудам (90 %) и ситовидным трубкам, а нисходящий – только по ситовидным трубкам. Лучше изучен восходящий транспорт. Для поступления воды в корень необходимо, чтобы водный потенциал ризодермиса был меньше, чем водный потенциал почвенного раствора; в этом случае вода начнет поступать в клетки.
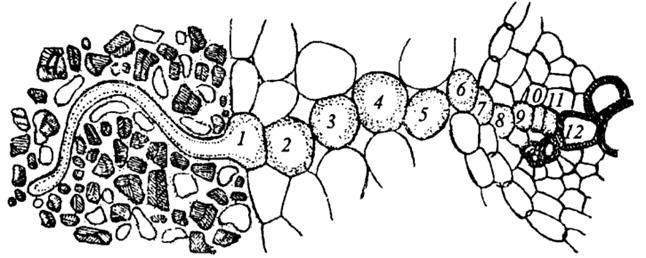
Путь воды от корневого волоска (1) до сосудов корня (12) через клетки паренхимы коры (2-6), эндодерму (7), перицикл (8) и паренхиму центрального цилиндра (9-11)
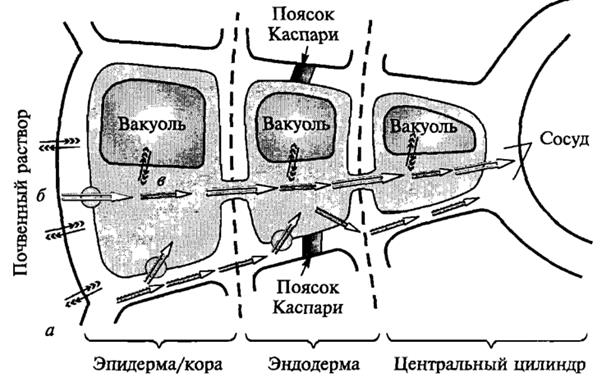
Итак, вода, попавшая в корневой волосок или другую клетку корня с помощью одного из механизмов, о которых мы говорили, движется к сосудам ксилемы, т.е. происходит радиальное перемещение воды по тканям корня. Движение воды в корневой волосок или другую клетку начинается с взаимодействия с оболочкой клеток, затем вода может пройти через плазмалемму и транспортироваться из протопласта одной клетки в протопласт другой через плазмодесмы. В этом случае вода движется по симпласту. Это симпластический путь. Однако вода могла бы и не заходить в симпласт, а оставаться в клеточной оболочке и двигаться по ней через ткани корня к центральному цилиндру. Это апопластический путь. Но по апопласту вода может двигаться только до эндодермы (до поясков Каспари). Поэтому путь воды по апопласту тут прекращается. Для дальнейшего передвижения она должна обязательно войти в симпласт. Необходимость переключения движения воды с апопластического на симпластический путь имеет большое значение, так как процесс движения по живой протоплазме клетка может сама регулировать – в отличие от перемещения воды по клеточным стенкам. Далее вода из клеток паренхимы центрального цилиндра корня поступает в сосуды ксилемы. Дальний транспорт. Путь, который проходит вода от корневого волоска до испаряющей клетки листа, распадается на две части, различные по протяженности, строению и физиологическим признакам. Первая часть состоит из живых клеток и имеет малые размеры (миллиметры или доли миллиметра). Это два коротких участка: один – в корне, от его поверхности с корневыми волосками до сосудов, которые находятся в его центральном цилиндре; второй – в листе, от сосудов, входящих в состав проводящего пучка, до испаряющей воду в межклетники хлоренхимы. Вторая часть пути – это сосуды, трахеиды, которые представляют собой мертвые трубки. У травянистых растений их длина составляет несколько сантиметров, у деревьев достигает нескольких метров и даже десятков метров. Вода и минеральные элементы доставляются к каждой клетке надземной части растения благодаря восходящему току по ксилеме. Существует нисходящий флоэмный ток растворов от листьев к корням. Направленный вниз флоэмный ток формируется в клетках мезофилла листьев, где часть воды, которая пришла с ксилемным током, с клеточных оболочек мезофилла переходит во флоэмные окончания. Вода из клеток листа и непосредственно из сосудов ксилемы поступает во флоэму по осмотическому градиенту, возникающему из–за накопленных в клетках флоэмы сахаров и других органических соединений, образующихся в процессе фотосинтеза. Нисходящий флоэмный ток доставляет органические вещества тканям корня, где они используются в метаболизме. В корнях окончания проводящих пучков элементов флоэмы, как и в листе, расположены вблизи элементов ксилемы, и вода вновь по осмотическому градиенту поступает в ксилему и движется вверх. Таким образом, происходит обмен воды в проводящей системе корней и листьев (как бы круговорот). Из–за движения тока воды по сосудам ксилемы при перерезании стебля какого–нибудь растения на небольшом расстоянии от почвы через некоторое время с конца сосудов начинает выделяться сок, который называют пасокой. Это явление получило название «плача растений», или экссудации. Силу, которая поднимает пасоку вверх по сосудам, назвали корневым давлением. Корневое давление можно измерить, если надетую на перерезанный стебель трубку соединить с манометром. Величина корневого давления непостоянна. В оптимальных условиях она составляет 2–3 бара. При определенных условиях достигается равновесие между количеством выделенной пасоки и количеством поступившей воды, поэтому корневое давление, или количество выделенной пасоки, может отражать поглотительную способность корней. Таким образом, активными двигателями начального восходящего водного тока (корневого давления) являются живые клетки, которые прилегают к нижнему концу проводящей системы растений – это клетки паренхимы корней – нижний концевой двигатель водного потока. Механизм корневого давления основывается на действии сократительных белков, функцию которых, как считают, выполняют микрофибриллы Ф–белков. В какой–то степени доказательством активного выхода воды может служить гуттация. Однако если бы растение постоянно не теряло воду в результате транспирации, то клетки корневых волосков быстро бы насытились водой и ее поступление прекратилось. Поэтому одной из причин возникновения градиента водного потенциала является испарение воды надземными органами. Чем интенсивнее клетки листьев испаряют воду, тем быстрее она начнет поступать в клетки корней и быстрее транспортироваться вверх по растению. Потеря молекул воды в верхней части водного столба в результате испарения заставляет воду течь по сосудам ксилемы вверх для ликвидации потери. Это вызванное транспирацией движение воды получило название транспирационного тока. Он обусловливает поступление воды из почвы в растение тоже по градиенту водного потенциала. Из–за транспирации водный потенциал в верхней части растения ниже, чем у основания. Активными двигателями водного тока, обусловленного транспирацией, являются живые клетки, прилегающие к верхнему концу всей проводящей системы растения – клетки паренхимы листа – верхний концевой двигатель водного тока. Механизм работы верхнего концевого двигателя несложный. Атмосфера обычно недонасыщена водяными парами, поэтому имеет отрицательный водный потенциал. При относительной влажности воздуха 90 % он составляет 140 бар. У большинства растений водный потенциал листьев колеблется от 1 до 30 бар. Из–за большой разности водных потенциалов происходит транспирация. Уменьшение количества воды в паренхимной клетке листа вызывает снижение активности воды в ней и уменьшение водного потенциала. Водный дефицит постепенно от клетки к клетке достигает корней, и активность воды в них снижается. В этом случае вода и поступает из почвы в корень. Таким образом, можно сделать вывод, что перемещение воды по растению, как и поступление ее в корень, связано с градиентом водного потенциала в системе «почва – растение – воздух». Если этот градиент начнет увеличиваться, тем больше воды станут терять клетки листа, т.е. чем сильнее транспирация, тем больше воды теряется. Работают два двигателя неодинаково, В среднем верхний концевой двигатель развивает силу в 10–15 бар и даже больше, а нижний, как отмечалось, 2–3 бара. Отсюда видно, что главная роль в водном обмене принадлежит верхнему двигателю. Однако при отсутствии листьев у деревьев зимой и ранней весной или после сухого периода основную роль в передвижении воды выполняет нижний двигатель. В поднятии воды по растению нижний концевой двигатель также участвует в условиях большей влажности воздуха, когда транспирация минимальна. Для верхнего концевого двигателя источник энергии – Солнце, это означает, что поглощаемая листом лучистая энергия используется для испарения. Для нижнего концевого двигателя источник энергии – дыхание. Энергия молекул АТФ, синтезируемых во время дыхания клеток корня, расходуется на транспорт ионов в клетке, т.е. на создание градиента водного потенциала. Таким образом, верхний концевой двигатель представляет собой автоматически работающий механизм, который тем сильнее присасывает воду, чем быстрее расходует ее. Работой верхнего и нижнего концевых двигателей без труда можно объяснить поднятие воды на несколько десятков сантиметров, пусть метров. А как объяснить поднятие воды на десятки метров (а секвойя достигает высоты 140 м)? Сосуды, по которым вода двигается на большей части своего пути, представляют собой мертвые трубки. Они не могут развивать силы для поднятия воды. Ответить на этот вопрос помогает теория сцепления, которую предложил английский исследователь Г. Диксон в 1921 г. В соответствии с этой теорией в сосудах образуются непрерывные нити, проходящие от клеток паренхимы корня до клеток паренхимы листьев. Сила, которая заставляет молекулы воды идти друг за другом, была названа силой сцепления (когезия). Непрерывные водяные нити образуются за счет водородных связей. Однако водяные нити сцеплены и со стенками сосудов (адгезия) с силой 300–350 бар. Все это позволяет нижнему и верхнему концевым двигателям поднимать воду по стволу на высоту 140 м. После появления этой теории анатомы не раз обращали внимание исследователей на образование пузырьков воздуха, которые должны нарушать сцепление между молекулами воды в сосудах. Однако в случае временного исключения какого–нибудь сосуда вода движется по запасным путям (другим сосудам) или апопласту, а воздушные пузырьки постепенно рассасываются при участии живых клеток. Передвижение воды из корня в лист по мертвым сосудам, оказывающим минимальное сопротивление водному току, – одна из находок природы. Клетки сосудов и трахеид вытянуты в длину, в них отсутствует живое содержимое, внутри они пустые, т.е. представляют собой пористые трубки. Одревесневшие вторичные клеточные оболочки достаточно крепки на разрыв, способны выдержать большую разность давлений, возникающих при подъеме воды к вершине больших деревьев. Торцевые, а иногда и боковые стенки члеников сосудов, перфорированы; сосуды, которые состоят из соединенных концами члеников, образуют длинные трубки, по которым вода с минеральными веществами легко проходит. В трахеидах нет перфораций, и вода, чтобы попасть из одной трахеиды в другую, должна пройти через их торцевые стенки; но трахеиды очень длинные клетки, а поэтому и это строение тоже хорошо приспособлено для проведения воды. Выход в процессе эволюции растений на сушу, крона которых располагается довольно далеко от земли, стал возможным благодаря образованию высокоспециализированной проводящей системы. Значение этого приспособления подчеркивается и названием самих растений – сосудистые. Кроме разницы в механизмах действия существует полная согласованность в работе двух концевых двигателей. На действие любого фактора среды, способного подавить работу нижнего двигателя, лист отвечает активацией транспирации, и наоборот. Это биологически важное приспособление, хотя, на первый взгляд, выглядит парадоксально: поступление воды ухудшается, а лист на этот неблагоприятный фактор отвечает не подавлением, а увеличением транспирации. Увеличение транспирации в данном случае имеет целью стимулировать поступление воды в корень. Сейчас существует взгляд, что в растении есть особая регуляторная система – гидродинамическая. Под ее контролем находятся водный режим, поддержание водного гомеостаза, а также некоторые другие функции, в частности фотосинтез. Гидродинамическая регуляторная система очень чувствительна. Она приходит в действие при очень маленькой потере воды листом (0,06 % от исходного количества) и предотвращает более сильное обезвоживание в дальнейшем. Передача сигнала к листу происходит через сплошной водный поток, а восприятие – устьичным и фотосинтетическим аппаратом. Рецептором сдвигов почвенных условий, тормозящих поступление воды, являются, скорее всего, мембраны эндодермальных клеток корня. Гидродинамическая регуляторная система позволяет растению очень быстро реагировать на внешние изменения, потенциально неблагоприятные для водного гомеостаза. Наземные растения стоят перед сложной дилеммой: они должны обладать развитой поверхностью, чтобы эффективно поглощать солнечный свет и СО2, но в этой связи увеличиваются потери воды. Эту проблему растения решают разными способами: во–первых, поступление воды увеличивается за счет роста корней и развития гипертрофированной поглотительной поверхности. Во–вторых, потеря воды становится медленнее из–за того, что мезофилльные клетки отделены от окружающей среды кутикулой, содержащей воск. В–третьих, противоречие между необходимостью поглощать большее количество СО2 и одновременно уменьшать количество испаряемой воды растения решают с помощью осцилляторного механизма, т.е. с 20–минутным интервалом последовательно происходят открытие и закрытие устьиц, в каждый промежуток времени интенсивность транспирации то увеличивается, то уменьшается. Дальнейший транспорт воды идет по проводящим тканям корня, стебля и листа. Опыты показали, что восходящий ток воды идет в основном по сосудам ксилемы, в которых она встречает незначительное сопротивление. Живые клетки оказывают большее сопротивление движению воды, поэтому по ним передвигается лишь 1–10 % ее общего количества. Из проводящих тканей стебля вода попадает в проводящие пучки листа. Чем больше проводящих пучков на единице листовой поверхности, чем сильнее они ветвятся, тем меньшее сопротивление встречает вода при своем движении в клетки мезофилла. Иногда мелких проводящих пучков так много, что они подводят воду почти к каждой клетке. Здесь вода из сосуда переходит в клеточные стенки, откуда испаряется в межклетники. Следовательно, в листовой пластинке транспорт воды идет в основном по апопласту. Транспорт воды в листовой пластинке изучен хуже, чем в корне. Большая часть водного пути приходится на долю проводящей системы растения, состоящей из сосудов и трахеид. У травянистых растений ее длина измеряется сантиметрами, а у деревьев – метрами.
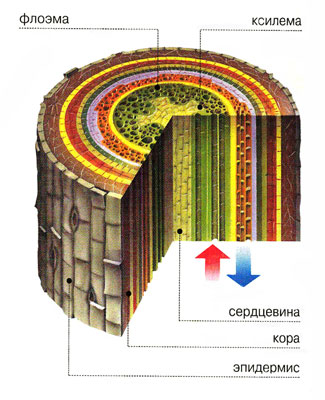
Ксилема выполняет в растении как опорную, так и проводящую функцию – по ней движутся вверх по растению вода и минеральные соли. В состав ксилемы входят элементы четырех типов: трахеиды, сосуды, паренхимные клетки и волокна. Трахеиды – мертвые одиночные клетки веретеновидной формы. Их концы перекрываются, придавая растению необходимую прочность. Вода движется по пустым просветам трахеид, не встречая на своем пути помех в виде клеточного содержимого; от одной трахеиды к другой она передается через поры. У покрытосеменных трахеиды развились в сосуды. Это очень длинные трубки, образовавшиеся в результате «состыковки» ряда клеток; остатки торцевых перегородок все еще сохраняются в сосудах в виде ободков. Размеры сосудов варьируют от нескольких сантиметров до нескольких метров. В первых по времени образования сосудах протоксилемы лигнин накапливается кольцами или по спирали. Это дает возможность сосуду продолжать растягиваться во время роста. В сосудах метаксилемы лигнин сосредоточен более плотно – это идеальный «водопровод», действующий на большие расстояния. Паренхимные клетки ксилемы образуют своеобразные лучи, соединяющие сердцевину с корой. Они проводят воду в радиальном направлении, запасают питательные вещества. Из других клеток паренхимы развиваются новые сосуды ксилемы. Наконец, древесинные волокна похожи на трахеиды. Они не проводят воду, но придают дополнительную прочность. Передвижение воды по сосудам ксилемы, длина которых может достигать нескольких десятков сантиметров, происходит довольно легко. Путь через трахеиды более трудный: от одной трахеиды к другой вода проходит через окаймленные поры; очевидно, что движение воды через них испытывает большее сопротивление, чем через сосуды. Измерения показывают, что у лиственных древесных растений проводимость древесины в 3 – 6 раз выше, чем у хвойных. В целом этот путь вода преодолевает куда легче, чем первый и третий через несколько миллиметров или даже долей миллиметров живых клеток – от корневых волосков до сосудов центрального цилиндра и от сосудов, расположенных в жилках листа, до испаряющих клеток мезофилла. По сосудам и трахеидам вода передвигается, как по полым трубкам, подчиняясь общим гидродинамическим законам, по живым же клеткам корня и листа – осмотическим путем, с помощью разности сосущих сил соседних клеток в правильно возрастающей последовательности. Значительное сопротивление току воды при переходе ее от одной живой клетки к другой делает этот способ совершенно непригодным для передвижения воды на большое расстояние. Поэтому возникновение трахеид у папоротниковидных растений явилось важным этапом в эволюции растительного мира. Еще более совершенной стала водопроводящая система с появлением настоящих сосудов у покрытосеменных растений. Через растение перекачивается огромное количество воды. С 1 га посевов пшеницы за лето испаряется около 2 тыс. т, клевера – 7,5, капусты – 8 тыс., Т.е. если собрать всю воду, расходуемую 1 га клевера или капусты, то получится водный бассейн площадью 1 га и глубиной 75– 80 см. Еловые молодняки южной тайги за год тратят 4,5 тыс. т воды с 1 га лесной площади, сосновые – 5,0, ольховые – до 11 тыс. т. Видно, что леса расходуют не меньше, а даже больше, чем некоторые сельскохозяйственные культуры. Эти колоссальные расходы восполняются за счет деятельности корневых систем, с достаточной быстротой всасывающих воду из почвы. На этом основана осушающая роль леса в условиях заболачивающихся лесных почв. Поддержанию водного баланса служат и хорошо развитая проводящая воду система, без задержки подающая воду к листьям, а также наличие покровных тканей, защищающих растение от излишней потери воды.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
 Установлено также, что вода и растворенные вещества могут передвигаться в стебле и в поперечном направлении. Это происходит, например, при подрезке корней или обрезке ветвей, в таком случае растение в состоянии обеспечить водой и веществами те части организма, которые раньше обслуживались утраченными органами. Возможность радиального транспорта обусловлена самой структурой проводящей системы, а именно: контактами сближающих трахеид через поры.
Установлено также, что вода и растворенные вещества могут передвигаться в стебле и в поперечном направлении. Это происходит, например, при подрезке корней или обрезке ветвей, в таком случае растение в состоянии обеспечить водой и веществами те части организма, которые раньше обслуживались утраченными органами. Возможность радиального транспорта обусловлена самой структурой проводящей системы, а именно: контактами сближающих трахеид через поры.