|
|
Физиология устьичных движенийУстьица открываются, когда в замыкающие клетки поступает вода, что приводит к увеличению тургорного давления в этих клетках и изменению их формы. Когда вода из замыкающихся клеток уходит, тургорное давление в них уменьшается, форма клеток изменяется и устьица закрываются. Существует несколько механизмов, влияющих на изменение тургорного давления в замыкающих клетках.
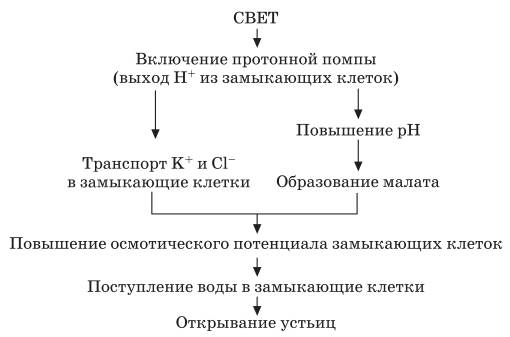
Схема открывания устьиц с участием осмотического механизма
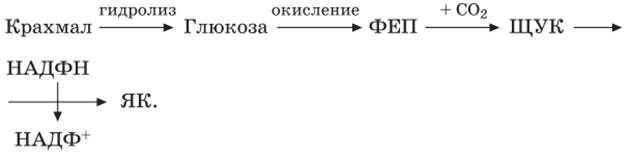
Больше всего тургор зависит от поступления ионов калия. При поступлении калия в замыкающие клетки в них уменьшается водный потенциал и вода поступает в клетки. При выходе ионов К+ увеличивается водный потенциал в замыкающих клетках и вода выделяется в свободное пространство клетки. Резервуаром К+ служат примыкающие клетки эпидермиса. Чтобы при поступлении ионов К+ в замыкающие клетки не изменялся мембранный потенциал, одновременно в эти клетки входят анионы или выходят ионы Н+. Источником протонов могут служить органические кислоты, поскольку при открытых устьицах их содержание в вакуолярном соке увеличивается. Так, яблочная кислота обеспечивает замыкающие клетки ионами Н* для обмена на К+ и анионами малата для баланса заряда наряду с ионами Cl-. Рассмотренные процессы могут быть представлены в виде схемы. Около 40 % ионов К+ входят в замыкающие клетки вместе с анионами Cl-. Доказательством выхода протонов из замыкающих клеток является повышение pH при поступлении в эти клетки ионов К+. Источником протонов могут служить органические кислоты, находящиеся в вакуолярном соке, поскольку при открытых устьицах их содержание увеличивается. Выход протонов приводит к подщелачиванию среды в замыкающих клетках, что вызывает гидролиз крахмала: Крахмал + Фн ↔ Глюкозо-1-фосфат. Реакция катализируется крахмальной фосфорилазой, чувствительной к концентрации водородных ионов. Поэтому поступление ионов калия в замыкающие клетки часто сопровождается распадом крахмала, а их выход – синтезом. Реакция обратима. Поэтому, когда образуется глюкозо-1-фосфат (осмотически активное вещество), водный потенциал уменьшается в замыкающих клетках и происходит поступление воды. В этом случае устьица открываются. Из-за увеличения концентрации глюкозо-1-фосфата осмотический потенциал может увеличиваться от 10–20 до 100 бар. Эта реакция превращения крахмала известна давно, а ее связь с движением устьиц получила название осмотического механизма движения устьиц.
Наибольшей чувствительностью к изменению pH в этой реакции обладает ФЕП-карбоксилаза. Некоторые авторы выделяют гидродинамический механизм движения устьиц. Он работает тогда, когда тем или иным путем вызывается остановка подачи воды в лист (перерезание черешка, опускание корней в растворы, тормозящие поступление воды в корень и передвижение ее по растению, и т.д.). Уже через несколько секунд после нарушения водного баланса листа устьичная щель увеличивается. Наличие хлоропластов в замыкающих клетках дает возможность говорить еще об одном механизме движений устьиц – фотосинтетическом. Когда концентрация СО2 в окружающей среде падает, восстановление его идет медленно создается возможность направить больше АТФ для работы протонных насосов, которые обусловливают поступление К+. Зависимость открытия и закрытия устьиц от расходования АТФ доказывает тот факт, что ингибиторы дыхания вызывают торможение движений устьиц. С другой стороны, ускорение восстановления СО2 приводит к образованию в замыкающих клетках сахаров, в результате уменьшается водный потенциал, вода поступает в клетки и устьица открываются. Регуляция функционирования устьиц обусловлена разными механизмами. Отмечается тесная связь во взаимодействии осмотического и фотосинтетического механизмов, т.е. одна и та же функция – движение устьиц – осуществляется с помощью разных взаимосвязанных механизмов. Другая физиологическая функция – поступление воды – также осуществляется с помощью различных механизмов: осмоса, набухания, электроосмоса. В этом один из интереснейших принципов организации физиологии клетки, ткани или органа вообще. Именно этот принцип следует назвать принципом надежности: когда выходит из строя один механизм, его дублирует другой, функция продолжает осуществляться. Это первая причина необходимости нескольких механизмов. Вторая состоит в следующем: из различных механизмов с помощью их комбинаций можно построить много разных систем, что ведет к лучшему приспособлению организма в окружающей среде. На ширину устьичных щелей наряду с парциальным давлением СО2 в межклетниках, общим содержанием воды в тканях, ионным балансом большое влияние оказывают и фитогормоны. Гибберелловая кислота и цитокинины способствуют открыванию устьиц. АБК, наоборот, приводит к закрытию, т.е. является антитранспирантом. Внешние факторы влияют на открывание устьиц, из которых наиболее значимы свет, температура, влажность воздуха, условия водообеспеченности. Действия эндо- и экзогенных факторов в ряде случаев взаимосвязаны. Поскольку ширина устьичных щелей зависит от тургесцентности замыкающих клеток, то вода – главный фактор, регулирующий движение устьиц. Свет влияет на движение устьиц через процесс фотосинтеза. Уменьшение концентрации СО2 увеличивает pH внутреннего содержимого замыкающих клеток, которое стимулирует разрушение крахмала и накопление глюкозо-1-фосфата. Поступает вода, и устьица открываются. Важным фактором является концентрация СО2. При определенной концентрации СО2 в воздухе устьица в темноте закрыты, но при снижении концентрации открываются. При этом не имеет значения, с какой стороны эпидермиса возникает нехватка СО2. Температура влияет, прежде всего, на скорость открывания устьиц. При температуре ниже 5 °С устьица открываются очень медленно и не полностью, а при отрицательных температурах (от 0 до -5 °С) остаются закрытыми. При температурах выше оптимальных они также открываются не полностью, но у некоторых видов ширина устьичных щелей при высокой температуре (40–45 °С) может снова достигать максимальной величины. Влияние температуры на движение устьиц происходит через влияние на скорость фотосинтеза и дыхания и на соотношение этих процессов. При высокой температуре интенсивность дыхания выше интенсивности фотосинтеза, в результате в межклетниках накапливается СО2 и устьица закрываются. При температурах, оптимальных для фотосинтеза, в межклетниках мало СО2 и устьица открыты. Малая интенсивность света, недостаток воды, высокая температура снижают интенсивность фотосинтеза и повышают интенсивность дыхания, увеличивают концентрацию СО2 в замыкающих клетках, что и вызывает закрывание устьиц. Высокая влажность около корней в почве, излишек калия способствуют открыванию устьиц. В регуляции функционирования устьиц участвуют прямые и обратные связи. Одни из них связаны с недостатком СО2, который может быть вызван фотосинтезом. Снижение межклеточной концентрации СО2 служит сигналом для открытия устьиц. Вода определяет другой тип связей, при ее недостатке в тканях устьица закрываются. Однако оптимизация газового обмена достигается прямыми связями – действием света, влажности и ветра и положительной связи фотосинтеза и обеспеченности водой эпидермальных клеток и (или) клеток мезофилла в значительной мере обусловлено синтезом АБК, а положительная обратная связь определяется продуктами фотосинтеза в клетках мезофилла. Внеустъичная регуляция транспирации представлена несколькими механизмами. Первый связан с обезвоживанием клеточных стенок, с поверхности которых идет испарение. Это механизм подсушивания. Когда поверхность верхних клеток мезофилла (хлоренхима) начинает подсыхать, то затрудняется движение Второй механизм связан со свойством протоплазмы изменять свою водоудерживающую способность. На его существование указывает то обстоятельство, что при одной и той же степени открытия устьиц транспирация может сильно меняться. Так, например, показано, что коротковолновые лучи (сине- и ультрафиолетовые) немедленно увеличивают транспирацию за счет изменения структуры белковых. При частичной денатурации способность белков удерживать воду ослабляется и кутикулярная транспирация возрастает. Толщина кутикулы на поверхности листьев влияет на интенсивность транспирации. Она может изменяться под влиянием внешних факторов. Увеличение толщины кутикулы уменьшает интенсивность транспирации с поверхности листьев. У некоторых растений существуют специфические механизмы внеустъичной регуляции транспирации. Так, например, у некоторых сортов сахарного тростника, когда транспирация превышает поступление воды, происходит свертывание листовых пластинок вдоль средней жилки в трубку и наблюдается снижение транспирации на 10–20 %. При нехватке воды растения могут терять листья. Опять же, у сахарного тростника может остаться на побеге в условиях засухи один лист. При возобновлении дождей или полива количество листьев быстро увеличивается. При наступлении зимней засухи в умеренной зоне деревья сбрасывают все листья. Однако листопадные формы деревьев встречаются в тропиках и пустынях.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|