
|
|
Поглощение воды растительной клеткойСОДЕРЖАНИЕ УЧЕБНОГО МАТЕРИАЛА Поглощение воды растительной клеткой.Физико-химические свойства воды и ее значение в жизни растений. Содержание и формы воды в растении. Клетка – осмотическая система. Осмотический, тургорный и водный потенциалы. Зависимость водного потенциала клетки от внутренних и внешних условий. Поглощение воды растением.Корневая система как орган поглощения воды. Особенности корневых систем древесных растений. Всасывающая и нагнетательная деятельность корней. Корневое давление и возможные механизмы его возникновения. Гуттация и «плач». Суточные и сезонные изменения корневого давления у древесных растений. Зависимость поглощения воды от внутренних и внешних условий. Связь водного обмена растений с водным режимом почвы. Характеристика форм почвенной влаги и их доступность растениям. Коэффициент завядания. Представление о «физиологической сухости» почвы. Транспирация.Лист как орган транспирации. Устьичная и кутикулярная транспирации. Физиологическое значение транспирации. Физиология устьичных движений. Регулирование транспирации. Количественные показатели транспирации. Методы измерения интенсивности транспирации. Зависимость транспирации от внешних условий и внутреннего состояния растений. Группы древесных растений по интенсивности транспирации. Испарение воды различными органами древесных растений. Условия, определяющие максимальную продуктивность транспирации. Суточная и сезонная динамика транспирации. Эвапотранспирация. Движение воды по растению.Путь водного потока в растении. Проводящие элементы ксилемы. Движущие силы водного потока. Поднятие воды от корней к листьям у древесных растений. Скорость передвижения воды по растению. Водопроводимость древесины хвойных и лиственных растений. Особенности движения воды в стволе дерева. Сезонные и суточные колебания содержания воды в древесном растении. Водный балансрастений.Уравнение водного баланса. Водный дефицит. Завядание растений. Связь водного обмена с другими физиологическими процессами и с продуктивностью. Регулирование водного обмена. Антитранспиранты. Поддержание водного баланса растения при пересадке растений. Тема 1 Поглощение воды растением 1. Поглощение воды растительной клеткой 2. Поглощение воды растением 3. Зависимость поглощения воды от внешних и внутренних факторов
Поглощение воды растительной клеткой Физико-химические свойства воды и ее значение в жизни растений. Среди всех веществ живой клетки вода занимает первое место, составляя 75 – 85 % ее массы. Исключением служат лишь клетки сухих семян и спор, обычно содержащих не более 14 % воды. Вода содержится не только в живых, но и в мертвых клетках (ксилема). В межклетниках она находится главным образом в виде пара. Нормальное функционирование клеток происходит при содержании воды равном 70 – 80 %. Вода обладает рядом специфических свойств, определяющих ее громадное значение в жизни растений. Она слабо диссоциирует на водород и гидроксилъный ион. Концентрация каждого из них составляет 10–7 г–ионов на 1 л воды. Чистая вода имеет поэтому значение рН 7. Вода имеет целый ряд особенностей, аномальных свойств по сравнению с близкими к ней по химическому составу веществами. Одним из аномальных свойств воды является ее плотность. Известно, что все вещества при нагревании увеличивают свой объем и в связи с этим уменьшают плотность. Исключение для воды состоит в том, что в интервале температуры от 0 °С до + 4 °С плотность ее не уменьшается, а увеличивается. В момент замерзанияобъем воды мгновенно увеличивается примерно на 11 %. Лед легче жидкой воды и держится на ее поверхности, плавает. В связи с этим вода в глубоких водоемах даже при очень сильных морозах не замерзает, ибо как только температура воды упадет ниже + 4 °С. она поднимается на поверхность и превращается в лед. У самого дна озера, например, температура воды зимой обычно равна + 4 °С, что весьма благоприятно для жизни его обитателей. Но вода может оставаться в жидком состоянии при температуре вплоть до –70 °С. Это переохлажденная вода. Способность воды сохраняться в переохлажденном состоянии в тканях растений, наличие в них растворов органических и минеральных веществ, способствующих сохранению жидкого состояния, предохраняет растение от гибели при трескучих зимних морозах. Необыкновенно высоки у воды температура кипения и замерзания. По всем физическим законам вода должна бы кипеть при температуре –75°С, а не при + 100°С и замерзать при –90°С. Эти особенности объясняются наличием структуры воды. «Исключением из правил» являются и ее необыкновенно высокие теплоемкость, поверхностное натяжение, растворяющая способность, прочность на разрыв (когезия), что также имеет неоценимое значение в жизненных процессах растений. Для водных растений существенное значение имеет отсутствие у воды цвета. Высокая светопропускающая способность воды делает возможным осуществление растениями под водой процесса фотосинтеза. В молекуле воды (Н2О) распределение зарядов (двух положительных и двух отрицательных) асимметрично. В связи с тем, что атомы водорода в молекуле расположены по отношению к атому кислорода под тупым углом 104,5°, молекула воды, будучи электронейтральной, имеет на одной стороне отрицательный, а на другой – положительный заряды. Такие молекулы называют диполями. Наличие дипольного момента, равного произведению величины заряда на расстояние между полюсами, определяет ориентацию молекул воды вокруг ионов: у катионов молекулы воды располагаются к поверхности иона своими отрицательными концами – атомами кислорода, а вокруг анионов – положительными, т.е. атомами водорода. Связывание молекул воды в электрическом поле ионов получило название электростатической гидратации. Явления электростатической гидратации наблюдаются и около ионизированных химических групп ряда органических соединений. Это имеет значение в поддержании особого состояния цитоплазмы, ее устойчивости и вместе с тем – лабильности. Гидратация – взаимодействие воды с гидрофильными (притягивающими воду) веществами, приводящее к изменению ее свойств. Различают два вида гидратации. Присоединение (притягивание) диполей воды к заряженным частицам различных ионов минеральных солей и ионизированных групп. Образование водородных связей с полярными группами органических веществ (между водородом воды и атомами кислорода или азота). Подвижность, активность воды непосредственно зависят от ее состояния в клетке. С этой точки зрения в физиологии растений принято различать свободную и связанную воду. По существу, вся имеющаяся в клетке вода находится в связи с веществами, входящими в состав клеточной стенки, протоплазмы и вакуолярного сока. Различны лишь форма и характер (прочность) этой связи, что обусловлено особенностями входящих в состав клетки химических соединений, а также взаимодействием этих соединений. К категории связанной воды относят следующие фракции воды: а) связанную осмотически (удерживаемая осмотически активными веществами, т.е. гидратно-связанная). Частицы осмотически связанной воды являются наиболее активными участниками физиологических процессов. На содержание осмотически связанной воды влияет, соответственно, количество находящихся в клетке растворенных веществ; б) коллоидно-связанную (гидратно-связанная), которая включает интрамицеллярную воду, находящуюся внутри коллоидной системы (в том числе и иммобилизированную воду), и интермицеллярную воду (находится на поверхности коллоидов и между ними). Коллоидно-связанная вода наименее подвижна и составляет наиболее стабильную часть водных запасов клетки, обеспечивающую устойчивость последнего в неблагоприятных условиях существования. Содержание коллоидно-связанной воды зависит от содержания гидрофильных биоколлоидов и степени их гидратации, которая в свою очередь связана с их водоудерживающей силой; 3) гидрофобно-стабилизированную (главным образом в мембранах). Физиологи условно понимают под связанной водой ту, которая не замерзает при понижении температуры до –10 °С. Всякое связывание молекул воды (добавление растворенных веществ, гидрофобные взаимодействия и др.) уменьшает их энергию. Именно это лежит в основе снижения водного потенциала клетки по сравнению с чистой водой. Капиллярно–поглощенная (пространственно–стабилизированная) вода находится в клеточных стенках и сосудах проводящей системы. Это наиболее подвижная форма, которую относят к категории свободной воды. Свободной называют воду, сохранившую все или почти все свойства чистой воды. Свободная вода легко передвигается, вступает в различные биохимические реакции, испаряется в процессе транспирации и замерзает при низких температурах. Клеточные стенки содержат две фракции капиллярно–поглощенной воды: подвижную и малоподвижную. Малоподвижная вода находится в микрокапиллярных пространствах, а так же удерживается водородными связями на микрофибриллах целлюлозы. Подвижная фракция составляет значительную часть воды, находится в крупных капиллярных пространствах между микрофибриллами целлюлозы и легко перемещается. Первое место по концентрации воды в клетке (около 98 %) занимает вакуоль. В вакуолярном соке соли, сахара и органические кислоты находятся в растворе, а белки, таннины и слизи составляют гидрофильную коллоидную фракцию вакуолей. В вакуолярном клеточном соке преобладает вода, удерживаемая сравнительно низкомолекулярными соединениями (осмотически-связанная) и свободная вода. Содержание воды в цитоплазме может достигать 95 % от массы цитоплазмы. Основной вид гидрофильных коллоидов в цитоплазме – белки. Вода, находящаяся на расстоянии до 1 нм от поверхности белковой молекулы, связана прочно (коллоидно-связанная вода). В самой цитоплазме имеется вода свободная, коллоидно- и осмотически-связанная. Кроме того, в протоплазме имеется определенное количество ионов, а следовательно, часть воды осмотически связана. Пластиды, митохондрии и ядра отделены от цитоплазмы собственными мембранами. Их объем изменяется под действием осмотических сил. Содержание воды в хлоропластах и митохондриях обычно меньше, чем в цитоплазме (в хлоропластах около 50 %), что связано с присутствием в них большого количества липидов и липофильных веществ. Наименьшее содержание воды характерно для клеточных стенок (30–50 %). Клеточные стенки обладают значительной гигроскопичностью благодаря в основном высокой гидрофильности их пектиновых и целлюлозных компонентов. В связи с этим в оболочке растительной клетки вода главным образом коллоидно-связанная. Передвижение воды вне проводящих пучков происходит главным образом по клеточным стенкам. Вода в растениях находится во всех органах и тканях, образуя непрерывную фазу. Высокое содержание воды характерно для вегетативных органов зеленых растений. В ходе индивидуального развития органа обычно наблюдается снижение водоудерживающей способности коллоидов протоплазмы, что приводит к уменьшению содержания воды в тканях. Низкое содержание воды характерно для семян, спор грибов. Основные функции воды в биологических объектах: 1. Водная среда объединяет все части организма в единое целое. В теле растения водная фаза представляет собой непрерывную среду на всем протяжении от влаги, извлекаемой корнями из почвы, до поверхности раздела жидкость – газ в листьях, где она испаряется. 2. Вода – важнейший растворитель и среда для биохимических реакций. 3. Вода участвует в упорядочении структур в клетках. Например, она входит в состав молекул белков, определяя их конформацию. 4. Вода – метаболит и непосредственный компонент биохимических процессов. Например, в фотосинтезе вода является донором электронов, в цикле Кребса она принимает участие в окислительных процессах. Вода необходима для гидролиза и для многих синтетических процессов. 5. Вода – главный компонент в транспортной системе высших растений при перемещении веществ по симпласту и апопласту. 6. Вода – терморегулирующий фактор. Она защищает ткани от резких колебаний температуры благодаря высокой теплоемкости и большой удельной теплоте парообразования. 7. Вода – хороший амортизатор при механических воздействиях на организм. 8. Благодаря явлениям осмоса и тургора (напряжения) вода обеспечивает упругое состояние клеток и тканей растительных организмов. Поглощение воды клеткой.Вода должна пройти сквозь две мембраны (плазмалемму и тонопласт) и через лежащую между ними цитоплазму (мезоплазму). Обычно все три структуры рассматриваются как единый мембранный барьер. Чтобы представить, каким образом вода проходит через мембрану, поместим клетку, в вакуоли которой находятся соли, сахара, аминокислоты и т.д., в сосуд с дистиллированной водой. Согласно молекулярно–кинетической теории, молекулы всех веществ находятся в состоянии быстрого хаотического движения, скорость которого зависит от энергии этих молекул. Поскольку молекулы воды малы и проходят через клеточные мембраны быстрее, чем молекулы других веществ, рассмотрим перемещение только молекул воды. Молекулы эти диффундируют во всех направлениях: в клетку и из клетки, в различные клеточные органеллы и из них. Вакуоль содержит значительные количества растворенных веществ. Молекулы этих веществ ослабляют связи между содержащимися в вакуоли молекулами воды, притягивая их к себе и тем самым уменьшая ее суммарный поток из клетки наружу. Растворенные вещества снижают активность молекул воды. Как следствие этого, кинетическая энергия воды в вакуоли ниже, чем в относительно более чистой воде вне клетки, т.е. снаружи вакуоли о любой участок ее мембраны ударяется в единицу времени больше молекул воды и большее их число проникает на этом участке внутрь, нежели из нее выходит. В результате этой быстрой неравномерной двусторонней диффузии молекул воды через мембрану тонопласта объем вакуоли увеличивается и создается тургор, содержимое клетки прижимается к ее стенке. Р = iCRT, где С – концентрация раствора в молях; Т – абсолютная температура; R – газовая постоянная; i – изотонический коэффициент (равный 1 + а(n – 1), где а – степень ионизации; n – количество ионов, на которое диссоциирует молекула электролита) Осмотический механизм поступления воды. Причиной односторонней диффузии является разность концентраций раствора по обеим сторонам мембраны. Система, которая содержит растворы разных концентраций (или раствор и растворенное вещество), разделенная мембраной, получила название осмотической. Пространство, окруженное избирательно проницаемой мембраной и заполненное каким–либо водным раствором, называется осмотической ячейкой. Клетка представляет собой осмотическую систему: более концентрированный раствор – это внутриклеточное содержимое (вакуолярный сок), менее концентрированный – раствор в свободном пространстве клетки, роль проницаемой мембраны, разделяющей эти пространства, играют плазмалемма, мезоплазма и тоноплает. Диффузия воды через избирательно проницаемую мембрану называется осмосом; концентрация растворенных веществ в вакуоли служит мерой максимальной способности клетки поглощать воду. Осмотическое давление можно выявить только в том случае, если раствор помещен в сосуд с избирательно проницаемой мембраной, по другую сторону которой находится растворитель. Таким образом, состояние осмотического давления в отсутствие проницаемой мембраны как бы потенциально, и поэтому ему дали название «осмотический потенциал» и обозначили буквой Р, Установлено, что осмотический потенциал пропорционален температуре и концентрации вакуолярного сока. Следовательно, величина Р не является постоянной. Для расчета потенциального давления закон Бойля–Мариотта для газов применяют формулу: Р = iCRT, где С – концентрация раствора в молях; Т – абсолютная температура; R – газовая постоянная; i – изотонический коэффициент (равный 1 + а(n – 1), где а – степень ионизации; n – количество ионов, на которое диссоциирует молекула электролита). Это выражение справедливо для разбавленных растворов и означает, что осмотическое давление при постоянной температуре определяется концентрацией частиц (молекул, ионов) растворимого вещества (количеством в единице объема раствора). Потенциальное осмотическое давление отражает максимально возможное давление раствора данной концентрации или максимальную способность раствора в ячейке поглощать воду. Таким образом, на величину осмотического потенциала клетки влияет концентрация веществ, растворенных в вакуолярном соке. Эти вещества называются осмотически активными. К ним относятся органические кислоты, аминокислоты, сахара, соли. Суммарная концентрация этих веществ в вакуолярном соке варьирует от 0,2 до 0,8 М. Величина осмотического потенциала разная у разных видов растений. Наименьшее осмотическое давление у растений, произрастающих в пресной воде (1–3 атм), у морских водорослей оно доходит до 36–55 атм. Для наземных однолетних растений характерна следующая закономерность: в более сухих местах растения имеют и большие значения осмотического потенциала. В условиях нормального водообеспечения осмотический потенциал клеток составляет 5–10 атм, на засоленных почвах – 60–80 атм и даже 100. Наибольший осмотический потенциал, который удалось отметить на засоленных почвах – 202,5 атм. Исключение составляют суккуленты. Вырастая на сухих местах, они имеют маленький осмотический потенциал, так как накапливают воду в клетках. Величина осмотического потенциала изменяется и в пределах одного растения: наименьшая – в корнях (5–10 атм), самая высокая – в верхних листьях (до 40 атм). У молодых растений осмотический потенциал меньше, чем у старых. Величина осмотического потенциала изменяется и в зависимости от внешних условий: обеспеченности водой, температуры, интенсивности света. Эти факторы определяют ее временные изменения. В полдень потеря воды в результате транспирации и накопления ассимилятов в клетках листьев вызывает увеличение осмотического потенциала. При хорошем водообеспечении, в частности у водных растений, колебания осмотического потенциала зависят только от скорости процесса фотосинтеза, связанной с изменением интенсивности света на протяжении суток. В меристематических клетках, не содержащих центральной вакуоли, также происходит осмотическое поступление воды, при этом избирательно проницаемой мембраной служит плазмалемма, а осмотически действующим раствором – цитоплазма. После работ В. Пфеффера поступление воды в клетку начали объяснять разностью осмотических потенциалов вакуолярного сока и наружного раствора. Когда клетка находилась в гипотоническом растворе, вода входила в нее (эндоосмос). Если же клетка помещалась в гипертонический раствор, тогда вода выходила из нее (экзоосмос). В последнем случае протопласт, вслед за сжимающейся вакуолью, мог отстать от клеточной оболочки, и тогда наблюдался плазмолиз. При одинаковых осмотических потенциалах растворов в клетке и снаружи (изотонический раствор) количество воды в клетке не меняется. Это объяснение поступления воды в клетку долгое время считали единственным. Однако в 1918 г. А. Уршпрунг и Г. Блюм (Германия) показали, что поступление воды в клетку зависит не только от разности осмотических потенциалов, а еще и от так называемой сосущей силы. Когда вода поступает в клетку, то увеличивается объем вакуоли и последняя оказывает все большее давление на цитоплазму. Цитоплазма прижимается к клеточной оболочке. Клеточная оболочка растягивается, и объем клетки увеличивается. Давление протопласта на клеточную оболочку получило название тургорного давления. Его стали обозначать буквой Т. Если бы клеточная стенка могла беспредельно растягиваться, тогда поглощение воды шло бы до уравнивания концентрации растворов снаружи и внутри. Но клеточная оболочка имеет небольшую эластичность, поэтому она давит на протопласт в противоположном направлении. Противодавление клеточной стенки на протопласт называется тургорным натяжением. Поскольку давление одинаково с противодавлением, то тургорное давление по абсолютной величине одинаково с тургорным натяжением. Являясь гидростатическим, тургорное давление противодействует дальнейшему поступлению воды в клетку. При поступлении воды в клетку давление клеточной оболочки на протопласт увеличивается и, в конце концов, становится равным осмотическому давлению. Поступление воды в клетку прекратится, наступит равновесие, состояние насыщения: количество воды в вакуоли не меняется, хотя молекулы воды продолжают перемещаться через мембрану в обоих направлениях. В состоянии насыщения осмотический потенциал равен по абсолютной величине тургорному давлению: Р = Т. Таким образом, в клетку может поступать вода только в том случае, если осмотический потенциал превышает тургорное давление. Сила S, с которой вода поступает в вакуоль, и была названа сосущей: S = Р – Т. Это выражение стало основным при определении величины поступления воды в клетку за счет осмотических сил.
Когда клетка находится в гипертоническом растворе, то при постепенной потере воды наступает плазмолиз. Внешний раствор легко проходит через клеточную оболочку и заполняет пространство между клеточной оболочкой и сокращающимся протопластом. При потере воды в результате испарения в воздушную среду благодаря большим силам сцепления между молекулами воды и клеточной стенкой протопласт, сокращаясь в объеме, не отстает от клеточной стенки, а тащит ее за собой. Клеточная оболочка, двигаясь за сокращающимся протопластом, выгибается и не надавливает на него, а наоборот, старается растянуть. В этом случае величина тургорного давления из положительной становится отрицательной, откуда S = Р – (–Т) = Р + Т. Это означает, что в условиях сильного обезвоживания сосущая сила клетки может быть больше, чем осмотический потенциал. Это состояние называется циторризом. По мере поглощения воды клеткой уменьшается осмотический потенциал из–за снижения концентрации вакуолярного сока, резко увеличивается тургор; в результате уменьшается сосущая сила. Если вода продолжает поступать в клетку, то сосущая сила уменьшается и становится равной нулю. Поступление воды прекращается. Следовательно, поступление воды за счет осмотических сил создает условия для ликвидации дальнейшего поглощения воды клеткой. Но клетка продолжает испарять воду, тургор уменьшается, вновь возникает сосущая сила. Таким образом, процесс поступления воды в клетку – саморегулируемый. Клетка может увеличить свой осмотический потенциал, таким образом, и сосущую силу с помощью активного транспорта в вакуоль органических и минеральных веществ, а также в результате ферментативного превращения нерастворимых веществ в растворимые (крахмал в сахар, белков в аминокислоты) или в результате распада гексоз на триозы, дисахаров – на моносахара. Специалисты по водному обмену растений Р. Слэтчер и С. Тейлор предложили вместо старого и широко используемого понятия «сосущая сила» использовать термин «водный потенциал». Как уже отмечалось, растворимые вещества понижают активность молекул воды, находящихся в клетке. Как результат этого, кинетическая энергия молекул воды в вакуоли ниже, чем в чистой воде снаружи, в частности в свободном пространстве. Энергетический уровень молекул вещества, который характеризуется скоростью их диффузии, получил название химического потенциала. Химический потенциал воды (μw) выражает максимальное количество внутренней энергии молекул воды, которое может быть превращено в работу, т.е. означает количество свободной энергии (размерность ккал/моль или Дж/моль):
где μw0 – химический потенциал чистой воды (принят равным нулю); aw – активность молекул воды; множитель RT необходим, чтобы перевести активность в единицы энергии. Активность чистой воды равна единице, а внутри клетки в растворах aw меньше единицы, поэтому величина lnaw отрицательная. Таким образом, химический потенциал воды в растворах и в клетке меньше, чем в чистой воде. Водный потенциал выражает способность воды в растворе произвести работу в сравнении с работой, которую производит чистая вода в этих же условиях:
где Водный потенциал характеризует способность воды диффундировать, испаряться или поглощаться. Он имеет размерность энергии, поделенной на объем (что совпадает с размерностью давления), и его величину выражают в атмосферах или барах (1 атм = 1,013 бар = 10 Па). В действительности мы не можем измерить энергию молекул воды, например в стакане, поэтому водный потенциал чистой воды взят за ноль при стандартных величинах температуры и давления. Измерить можно только разность энергий молекул воды, находящихся в разных условиях. Чем ниже энергия молекул воды, тем ниже и водный потенциал. Поскольку водный потенциал чистой воды принимается равным нулю, то при увеличении концентрации растворенных веществ он становится более отрицательным. Водный потенциал раствора меньше, чем чистой воды, поэтому молекулы воды в вакуоли обладают меньшей свободной энергией, меньшим потенциалом, чем находящиеся снаружи клетки. В соответствии со вторым законом термодинамики процессы переноса веществ и энергии самопроизвольно идут от более высокого уровня химического потенциала к более низкому, т. е. по градиенту потенциала. Таким образом, вода движется в соответствии со вторым законом термодинамики из области высокого потенциала в область низкого, т. е. направление потока воды определяется падением градиента энергии. Поэтому правильнее говорить, что вода не поглощается клеткой, а поступает в клетку за счет разницы водных потенциалов. В физиологии растений часто используют понятие «матричный потенциал». Он характеризует снижение активности воды за счет гидратации коллоидных веществ и адсорбции на границе раздела фаз. Когда молекулы воды ассоциированы с поверхностями раздела, которые образуются, например, при суспендировании коллоидных частиц в водном растворе, они обнаруживают меньшую тенденцию реагировать химически в основной массе раствора и испаряться в окружающую газовую фазу. Это происходит потому, что вода имеет конечную константу связывания на поверхности раздела «жидкость – твердое тело», в результате молекулы воды постоянно подходят и уходят с таких поверхностей. Это приводит к связыванию части молекул воды и, следовательно, понижает термодинамическую активность воды aw, особенно вблизи коллоидов. Присутствие растворенных веществ также понижает активность воды. Матричные потенциалы используются и для описания химического потенциала воды в почве и в клеточных оболочках. Водный потенциал клетки, ткани или органа, целого растения – величина интегральная. Он состоит из осмотического Ψр, матричного Ψm, тургорного ΨT и гравитационного Ψg потенциалов:
Осмотический потенциал обусловлен наличием в клетках осмотически активных веществ и отражает их влияние на активность воды. Величина его равна величине осмотического давления с обратным знаком. Величина матричного потенциала определяется силами адсорбции между биополимерами цитоплазмы и молекулами воды, т.е. Ψm отражает влияние макромолекул полимеров на активность воды. Тургорный потенциал обусловлен эластичным противодействием клеточной оболочки разбуханию и характеризует влияние на активность воды тургорного (гидростатического) давления. Гравитационный потенциал отражает влияние на активность воды сил притяжения только при поднятии воды на значительную высоту. Величины Ψp Ψm и Ψg всегда отрицательные, так как присутствие растворенных веществ, биополимеров, а также действие сил тяжести снижают активность воды. ΨT, наоборот, положительный, поскольку при действии на воду механического давления (тургорного или гидрастатического) активность молекул воды увеличивается. Поскольку в большинстве случаев величины как матричного, так и гравитационного потенциалов малы (травянистые растения), то Ψw = Ψp + ΨT, а водный потенциал клетки будет равен Ψкл = –Ψm – Ψm т, при Ψр = ΨТ клетка не будет поглощать воду (Р = Т). Таким образом, если клетку опустить в воду, то она станет поглощать воду до тех пор, пока гидрастатическое давление не сравняется с осмотическим потенциалом, т.е. пока водный потенциал клетки не станет равным нулю. После этого в клетку уже не будет поступать вода ни из какого раствора или из другой клетки. Если рядом расположены две клетки с разными водными потенциалами, то вода через клеточную оболочку начнет проходить из клетки с более высоким водным потенциалом (менее отрицательным) в клетку с более низким (более отрицательным). Когда влажность почвы достаточная, а испарение не очень интенсивное, клеточная оболочка насыщена водой. В этом случае водный потенциал клеточной оболочки выше, чем в вакуоли, и вода поступает в клетку, в вакуоль. При недостатке воды в почве в клеточной оболочке может наблюдаться водный дефицит и, таким образом, водный потенциал будет ниже, чем в вакуоли, в результате вода станет из вакуоли вытекать. В последние годы установлено, что транспорт воды осуществляют специальные белки – аквапорины, формирующие в мембране селективные для воды каналы. Название «аквапорины» произошло от лат. agua – вода и греч. poros – отверстие. Молекула такого белка состоит из шести субъединиц, пересекающих липидный бислой. Внутренняя поверхность поры гидрофильна, что позволяет молекулам воды легко проникать через мембрану.
Образованные аквапоринами поры (каналы) ускоряют диффузию молекул воды. Помимо переноса воды через мембрану они транспортируют малые молекулы органических веществ, обеспечивающих осморегуляцию в клетке. Транспортная активность аквапоринов регулируется их фосфорилированием/дефосфорилированием, т.е. регуляция мембранной проницаемости для молекулы воды осуществляется посредством присоединения или отщепления остатка фосфорной кислоты в молекуле аквапорина. Этим путем растительные клетки регулируют проницаемость мембран для молекул воды. Такой тип изменения активности аквапоринов влияет не только на скорость транспорта, но и на направление водного потока. Коллоидно–химический механизм (набухание) поступления воды. В клетку вода может поступать и в результате набухания. Набуханием называют поглощение жидкости или пара высокомолекулярным веществом (набухающим телом), сопровождаемое увеличением объема. Явление набухания обусловлено коллоидными и капиллярными эффектами. В протоплазме наблюдается набухание на коллоидной основе (гидратация протоплазматических коллоидов), а в клеточной стенке – за счет капиллярного (накопление воды между микрофибриллами и в межмицеллярных пространствах) и коллоидного (гидратация полисахаридов, особенно гемицеллюлоз) эффектов; в вакуоли, как правило, набухающих тел нет. Состояние набухания протоплазмы имеет решающее значение для интенсивности всего обмена веществ. Вода в протоплазме – важнейшая среда для биохимических реакций и диффузии, а гидратация протоплазматических белков необходима для поддержания ультраструктуры и функциональной активности органоидов. У некоторых частей растения поступление воды происходит исключительно путем набухания, например у семян и у многочисленных приспособлений эпифитов для поглощения воды и водяного пара (всасывающие чешуйки и др.). Вода проникает в набухающее тело путем диффузии. Удерживаемые набухающим телом молекулы воды теряют часть своей кинетической энергии, которая превращается в тепло; теплоту набухания можно измерить (например, энергию набухания семян измеряют в закрытом термосе со вставленным термометром). Благодаря большому сродству набухающего тела к воде при набухании может возникнуть давление в несколько сотен атмосфер (например, разрыв гипсовых блоков набухающими семенами). Изменение конфигурации белковых молекул может привести к ситуации, когда водородные связи разрушаются, в этом случае вода может выталкиваться клеткой даже тогда, когда окружающий воздух насыщен водой. Это наблюдается в период поспевания семян, в период их обезвоживания. Например, в коробочках мака при большой концентрации в полостях водяных паров происходит обезвоживание семян путем изменения водоудерживающей способности клеток семян. Между вакуолью, цитоплазмой и клеточной оболочкой после каждого изменения в содержании воды сразу устанавливается равновесие. Электроосмотический механизм. Кроме двух названных механизмов существует еще третий механизм – электроосмос. Это понятие ввели Т. Беннет–Кларк с сотрудниками (1946). Электроосмос – перемещение диполей воды, вызванное электрическим потенциалом, возникающим на мембране. Разность электрических потенциалов (РП) на мембране – результат асимметричного распределения ионов по двум ее сторонам. Эта разность потенциалов, измеряемая десятками милливольт, может Однако мы имеем мало доказательств существования этого механизма. Одно из них – опыт Т. Беннета–Кларка с сотрудниками. Клетки эпидермиса лука помещали в раствор сахарозы и регистрировали объем протопласта. Затем раствор сахарозы заменяли изотоническим раствором КС1, и отмечалось уменьшение объема протопласта. Когда ткань вновь помещали в изотонический КС1 раствор сахарозы, объем протопласта увеличивался. Это объяснили следующим образом. Повышение концентрации К+ во внешней среде уменьшало РП на мембране. Уменьшение РП нарушает электроосмос, и объем протопласта уменьшается. Вероятно, существует три механизма, обусловливающих поступление воды в растительную клетку: 1. Поступление за счет осмоса, где главную роль играет вакуоль. 2. Поглощение воды биоколлоидами протоплазмы и структурными элементами клеточной оболочки (набухание). 3. Поступление за счет электроосмоса, обусловленного РП на мембране. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|

 Если клетка полностью насыщена водой (тургенесцентна), S = 0, тогда Р = Т. Когда подача воды уменьшается (ветер, недостаток влажности в почве и т.д.), то возникает водный дефицит в клеточных оболочках, водный потенциал в них уменьшается (становится ниже, чем в вакуолях), вода начинает передвигаться в клеточные оболочки. Отток воды из вакуоли снижает тургорное давление, и, таким образом, увеличивается сосущая сила. При длительном недостатке влаги большинство клеток теряет тургор. В этих условиях T = 0, S = Р.
Если клетка полностью насыщена водой (тургенесцентна), S = 0, тогда Р = Т. Когда подача воды уменьшается (ветер, недостаток влажности в почве и т.д.), то возникает водный дефицит в клеточных оболочках, водный потенциал в них уменьшается (становится ниже, чем в вакуолях), вода начинает передвигаться в клеточные оболочки. Отток воды из вакуоли снижает тургорное давление, и, таким образом, увеличивается сосущая сила. При длительном недостатке влаги большинство клеток теряет тургор. В этих условиях T = 0, S = Р.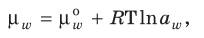
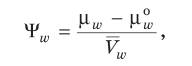
 – парциальный молярный объем воды, т.е. объем 1 моля воды (18,0 см3/моль).
– парциальный молярный объем воды, т.е. объем 1 моля воды (18,0 см3/моль).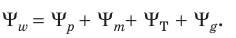
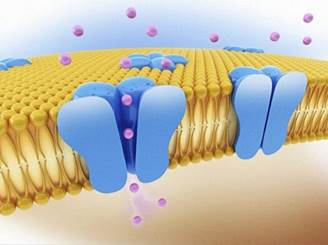 Аквапорины найдены в клетках практически всех растений. Особенность аквапоринов в их разнообразии. Например, в растениях хрустальной травки обнаружено более 30 генов, кодирующих белки водных каналов. Все аквапорины делят по месту их локализации в клетке на две группы: аквапорины плазмалеммы и аквапорины тонопласта.
Аквапорины найдены в клетках практически всех растений. Особенность аквапоринов в их разнообразии. Например, в растениях хрустальной травки обнаружено более 30 генов, кодирующих белки водных каналов. Все аквапорины делят по месту их локализации в клетке на две группы: аквапорины плазмалеммы и аквапорины тонопласта.