|
|
Поглощение воды растениемДля нормального функционирования клетки растения должны быть насыщены водой. Состояние насыщения поддерживается с помощью двух взаимосвязанных процессов: поступления и выделения (расходования) воды, которые и составляют водный обмен растений. Соотношение между этими процессами называют водным балансом. При анализе соотношения между поступлением и расходованием воды возможны три случая: поступление больше расходования, равно или меньше. В последнем случае возникает водный дефицит. В полдень водный дефицит может достигать 5–10 и даже 25 %. Одно из условий нормального функционирования наземных растений – поддержание условий без длительного и глубокого водного дефицита. Для этого необходима хорошо развитая корневая система, обеспечивающая поступление воды с большой скоростью. Поступление воды из почвы в корень. Поступление воды из почвы – более сложный процесс, чем простое всасывание воды растением, опущенным в водный раствор корнями. В почве мы имеем ряд сил, противодействующих этому всасыванию, которые можно назвать водоудерживающими силами. Перед тем как перейти к разговору о радиальном транспорте воды, рассмотрим в общих чертах строение поперечного среза корня.
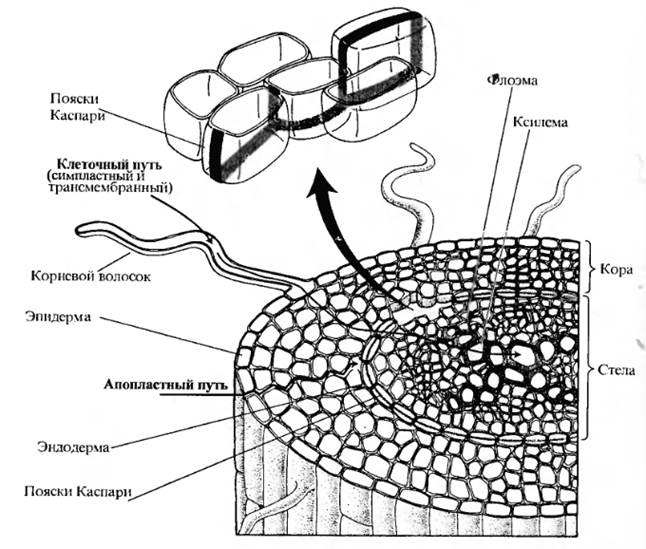
Эпиблема состоит из одного слоя клеток, покрывает молодые корни; ее главная функция – поглощение веществ. Наружные стенки клеток эпиблемы не имеют ни кутикулы, ни воска, поэтому они хорошо приспособлены для поступления воды. Некоторые летки эпиблемы вытягиваются и превращаются в корневые волоски. Следовательно, эпиблема как поглощающая ткань неоднородна. Благодаря корневым волоскам поверхность корня, соприкасающаяся с почвой, увеличивается в 10–15 раз. Кроме того, транспортные белки, находящиеся в плазмалемме корневых волосков, активнее, чем в других клетках; из корневых волосков идет больше плазмодесм в клетки экзодермы. Корневые волоски живут несколько дней и разрушаются. Вместо них в верхней части зоны растяжения образуются новые. Корневой волосок имеет большую центральную вакуоль, поэтому главный механизм поступления в него воды – осмос. Несмотря на важную роль корневых волосков, у некоторых растений, например у авокадо, они отсутствуют. Их нет и у водных растений. Постепенно по мере роста корня эпиблема заменяется пробкой. Через опробковевшие части вода почти не поступает в корень. Большую часть (до 80–90 %) площади поперечного сечения молодого корня составляет первичная кора. В ее крупных клетках центральные вакуоли занимают около 90 % объема протопласта. В состав клеточных стенок входит только целлюлоза, поэтому они хорошо проницаемы для воды. В коре много межклетников, которые в зависимости от условий могут быть заполнены воздухом или водой. Чем толще слой коры, тем больше диаметр корня и, следовательно, площадь его поверхности, покрытая эпиблемой. Внутри корня находится центральный цилиндр, отделенный от коры эндодермой. Эндодерма состоит из одного слоя клеток, в радиальных и поперечных стенках которых откладывается суберин, а у некоторых растений – лигнин. Образуются так называемые пояски Каспари, не проницаемые для веществ и воды. Это влияет на транспорт последних внутри корня. Непосредственно за эндодермой находится перицикл. В состав центрального цилиндра входят проводящие ткани. Большие сосуды ксилемы окружены живыми клетками ксилемной паренхимы. Крупные поры в клеточных стенках соединяют их с сосудами. Флоэма состоит из ситовидных трубок, клеток-спутниц и флоэмной паренхимы. С помощью проводящих тканей поддерживается связь между корнем и другими частями растения. Все эти ткани постепенно образуются из меристематических клеток, находящихся на верхушке, поэтому по длине в корне различают 4 зоны: деления клеток, их растяжения, зону дифференцировки (зону корневых волосков) и зону опробковения. Клетки зоны деления защищены корневым чехликом. Вода поступает главным образом в клетки зон растяжения и особенно корневых волос- Корень обладает еще одним важным свойством – положительным гидротропизмом, т.е. при недостатке воды растущие части корня изгибаются и растут в сторону влажных участков почвы. Для поступления воды в корень необходимо, чтобы водный потенциал ризодермиса был меньше, чем водный потенциал почвенного раствора; в этом случае вода начнет поступать в клетки. Итак, вода, попавшая в корневой волосок или другую клетку корня с помощью одного из механизмов, о которых мы говорили, движется к сосудам ксилемы, т.е. происходит радиальное перемещение воды по тканям корня. Все многообразие древесных растений по характеру строения и развития корневых систем делят на три основные группы: · с глубоко идущей в почву якорной корневой системой (лиственница, дуб, липа, робиния и др.); · с поверхностной корневой системой (ель, ясень, рябина и др.); · с промежуточным типом корневых систем, включающим в зависимости от лесорастительных условий как глубоко идущую, так и поверхностную корневую систему (сосна, береза, осина, многие клены, ольха и др.). Далеко не вся корневая система всасывает воду. Этой способностью обладают лишь молодые корневые окончания. Вода поглощается клетками зоны корневых волосков, а также зоны растяжения. Поглощающие корни относятся к последнему порядку ветвления. Для них характерно первичное строение, отсутствие камбиального слоя. Основная их функция поглощение воды и элементов минерального питания из почвы, тогда как для ростовых – увеличение размеров корневых систем. Длина всасывающей части корня травянистых растений достигает нескольких сантиметров, а древесных – всего лишь нескольких миллиметров. При этом у большинства деревьев всасывающий аппарат увеличивается за счет развития микоризы – симбиоза корня с гифами гриба. Грибной чехол микоризных корней и отходящие от него отдельные гифы существенно увеличивают поглощающую поверхность корня. Оказалось также, что микоризные растения более устойчивы к засухе, чем растения без микоризы: грибница продолжает активно всасывать воду при таком осмотическом давлении почвенного раствора, когда безмикоризные корни уже не работают. Различают пассивное и активное поглощение воды корневой системой. Первое осуществляется за счет сосущих сил листьев, теряющих воду в процессе транспирации. Активное поглощение тесно связано с метаболизмом клеток корня и проявляется в явлениях «плача» и гуттации. При перерезании стебля через поверхность среза начинает вытекать жидкость – пасока. Это явление носит название «плача» растений. Очень легко наблюдается «плач» у таких растений, как тыква, гортензия, а из древесных – у березы, кленов, особенно сахарного. В пасоке березы при весеннем сокодвижении содержится до 1,5 % сахаров, а у отдельных деревьев до 3,5 % (сахароза, глюкоза и фруктоза). В небольших количествах в березовом соке обнаруживаются аминокислоты, жирные кислоты, минеральные соли и некоторые другие вещества. Эти качества пасоки широко используют в народном хозяйстве. Она добывается в процесс е так называемой ксилемной подсочки берез, кленов и некоторых других пород. Под ксилемной подсочкой понимается совокупность производственных процессов, связанных с добычей ксилемного сока лиственных древесных растений. Добываемая жидкость (пасока) используется для приготовления сока, сиропа, кваса, вина, спирта, а также в качестве добавок в парфюмерно-косметическом производстве, медицине, животноводстве и пчеловодстве. Грамотное ведение ксилемной подсочки березы в 5 раз экологически более эффективно, чем сбыт березовой древесины с единицы лесопокрытой площади. Еще более выгодна пневая подсочка берез. Ксилемная подсочка лиственных древесных растений существенно снижает и затраты картофеля и других ценных с/х продуктов в спиртовом производстве. Иногда можно наблюдать выделение капелек жидкости из неповрежденных листьев. Это гуттация. Она ярко проявляется у проростков злаков, У каллы, манжетки, земляники. Из древесных растений нередко гуттируют ольха, ивы, вязы и некоторые другие. Гуттация обусловлена тем, что при высокой влажности воздуха и почвы в теплую погоду вода, поступившая в растение, не испаряется, а выдавливается в виде капелек жидкости. У срезанного побега дерева или кустарника даже при 100 % влажности воздуха гуттация наблюдаться не будет, ибо отсутствует корневое давление.
Гуттация происходит через специальные железки – гидатоды
«Плач» и гуттация связаны с наличием корневого давления, нагнетающего воду вверх из корней по растению. Само корневое давление является результатом жизнедеятельности корня. Движущей силой корневого давления служит энергия аэробного дыхания, ибо ингибиторы дыхания (яды, наркотики) и анаэробиоз приводят к прекращению корневого давления. Роль процесса дыхания в проявлении корневого давления может быть не только прямой, но и косвенной. Энергия этого процесса способствует поглощению ионов солей из почвы, передвижению их по живым клеткам корня и выделению в полости сосудов центрального цилиндра корня. При интенсивном дыхании концентрация солей в сосудах корня увеличивается, а вместе с ней повышается осмотическое давление раствора в сосудах. Существуют две основные гипотезы, объясняющие механизм действия корневого давления: Осмотuческая теория основана на явлениях осмоса. В сосудах центрального цилиндра поглощающего корня всегда находится раствор определенной концентрации, обладающий сосущей силой, равной целиком осмотическому давлению, так как в мертвых элементах ксилемы отсутствует тургорное давление. Поэтому вода будет отсасываться от ближайшей к сосуду клетки. Теряя воду, эта клетка будет развивать большую сосущую силу, что позволит ей отнимать воду от соседней с ней клетки и т.д. Клетки коровой паренхимы и центрального цилиндра корня будут испытывать недонасыщение водой. Этот дефицит коснется и корневого волоска, способного поглощать воду из почвы. При этом распределение величины осмотического давления в ряду клеток, про водящих воду, не играет никакой роли, ибо основным фактором осмотического поглощения воды выступает сосущая сила клеток. Метаболuческая теория основывается на допущении, что клетки корня, через которые наблюдается односторонний ток воды, обладают полярностью. На противоположных сторонах осуществляются диаметрально разные процессы: на одной (обращенной к периферии) синтезируются осмотически активные сахара, а на противоположной – инертный в осмотическом отношении крахмал. В первом случае осмотическое давление и сосущая сила будут больше, следовательно, к этой стороне клетки будет поступать вода от соседней клетки и выдавливаться с противоположного конца в сторону сосуда. Так создается непрерывный односторонний ток воды в корневой системе, как результат развиваемого в корнях корневого давления. В области эндодермы корня вода проходит через ее пропускные клетки, но их сравнительно мало. Другие клетки эндодермы с суберинизированными поясками Каспари, тесно связанными с плазмалеммой, представляют сильное препятствие току воды и растворов. По существу, эндодерма – основной барьер, препятствующий одностороннему току воды из клеток первичной коры в центральный цилиндр корня. Недавно с помощью современной техники было показано, что растворы проходят только через протопласты клеток эндодермы, тогда как в других клетках – как через симпласт, так и через апопласт (по клеточным стенкам). Величина корневого давления обычно колеблется около 1 атм, реже 2,5 - 3,0 атм. С помощью корневого давления растения в ночное время суток восполняют водный дефицит, возникающий в жаркий летний день. Корневое давление, у древесных растений действует в течение всего года, в том числе зимой, пополняя водные запасы ствола. Как установил Л. А. Иванов, после спиливания дерева влажность древесины пня повышается под действием корневого давления. Особенно велико значение корневого давления ранней весной до распускания листьев, когда отсутствует притягивающая воду транспирация. Корневое давление можно определить с помощью манометра, прикрепленного к пеньку срезанного растения. В обычных условиях роль пассивного поглощения воды в водообмене растений существенно выше. Оно более выгодно и в энергетическом отношении, так как не связано с затратой внутренней энергии, присущей активному поглощению, а осуществляется за счет энергии Солнца.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|