|
|
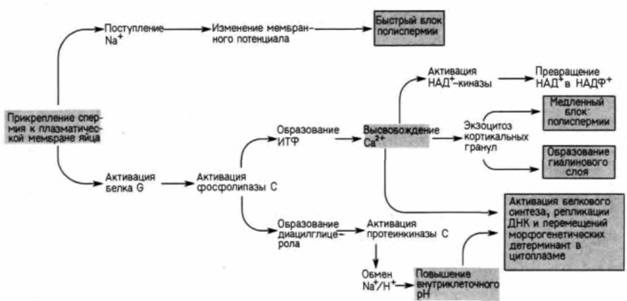
Активация метаболизма яйцаДо сих пор мы обсуждали механизмы, с помощью которых спермии и яйцо узнают друг друга, сливаются и благодаря которым происходит слияние их гаплоидных ядер. Но для того чтобы оплодотворение привело к развитию яйца, должны произойти изменения в его цитоплазме 1. Зрелое яйцо морского ежа представляет собой метаболически инертную клетку, которую активирует проникающий в нее спермий. Эта активация является, однако, только стимулом, запускающим запрограммированную цепь метаболических событий. Реакции яйца на воздействие спермия можно подразделить на «ранние», которые осуществляются в течение нескольких секунд после соединения гамет, и «поздние», происходящие спустя несколько минут после этого соединения (табл. 2.2). Ранние реакции Как мы видели, контакт между спермием и яйцом запускает два основных механизма блокирования полиспермии: быстрый блок, инициируемый поступлением в клетку натрия, и медленный блок, инициируемый повышением внутриклеточной концентрации ионов кальция. Активация яиц, по-видимому, всегда обусловлена повышением концентрации свободных ионов кальция в цитоплазме. У первичноротых, например у улиток и червей, кальций обычно поступает в яйцо из внешней среды. У вторичноротых, например у рыб, лягушек и морских ежей, активация осуществляется в результате выхода ионов кальция из внутриклеточных депо, причем возникает волна высвобождения ионов кальция, распространяющаяся по поверхности яйца (Jaffe, 1983). Результаты ряда опытов свидетельствуют о том. что этот выход ионов кальция играет важную роль в активации развития зародыша. Если в яйца инъецировать хелирующее кальций вещество ЭГТА,— то в них не происходит увеличения концентрации свободных ионов кальция и не наблюдается активации метаболизма. Напротив, яйца можно активировать искусственно, в отсутствие спермиев,
посредством воздействий, вызывающих выход свободного кальция в цитоплазму. Было показано (Steinhardl. Epel, 1974), что микромолярные количества кальциевого ионофора A23187 вызывают у большинства яиц реакции, характерные для оплодотворенных яиц: в нормальной последовательности происходят отделение оболочки оплодотворения, повышение внутриклеточного pH, скачкообразное увеличение потребления кислорода, интенсификация синтеза белков и ДНК. Существенно, что такая активация может происходить при полном отсутствии ионов кальция в морской воде. В большинстве случаев развитие таких искусственно активированных яиц останавливается на стадии первого деления дробления, так как они остаются гаплоидными и не содержат центриоли спермия, необходимой для деления. Выход кальция активирует целый ряд метаболических реакций (рис. 2.32). Одна из них заключается 1 У некоторых видов хвостатых амфибий функция оплодотворения, заключающаяся в побуждении яйца к развитию, полностью обособилась от его генетической функции. Серебристая амбистома (Ambystoma platineum) является видом, представленным только одними самками. Самки производят яйца с нередуцированным числом хромосом. Такие яйца, однако, неспособны к развитию без дополнительной стимуляции. Серебристые амбистомы спариваются с самцами амбистомы Джефферсона (A. jeffersonianum). Спермии этих самцов только побуждают яйца к развитию (Uzzell. 1964), не участвуя в формировании генома зародыша.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 64 _________ ГЛАВА 2
в увеличении потребления яйцом кислорода. Удивительно, что этот скачок в использовании кислорода происходит не в митохондриях и, возможно, мало связан (или совсем не связан) с образованием АТФ. Вероятнее всего, кислород используется для окисления различных компонентов яйца, в частности оболочки оплодотворения. Показано (Foerder et al., 1978). что эта оболочка, имеющая полужидкую консистенцию, затвердевает при окислении остатков тирозина, связанных с белками. Одновременно окисляются также разные липиды цитоплазмы (Perry. Epel. 1977). Другим кальций-зависимым эффектом является активация фермента НАД+-киназы, которая превращает HАД+ в НАДФ+ (Epel et al.. 1981). Это превращение может иметь важные последствия для метаболизма клетки: НАДФ+ (но не НАД+) используется в качестве кофермента при биосинтезе липидов. Таким образом, превращение НАД+ в НАДФ+ играет важную роль в создании многих новых компонентов плазматической мембраны, необходимых в процессе дробления. Поздние реакции Повышения содержания свободных ионов кальция недостаточно для осуществления всех реакций оплодотворения. Оно сочетается с повышением внутриклеточного pH. Полагают, что сочетание этих двух условий (высокое содержание Са+ и низкий pH) приводит к осуществлению полного спектра явлений оплодотворения, включая белковый синтез и синтез ДНК (Winkler et al.. 1980: Whitaker, Steinhardt. 1982). Повышение внутриклеточного pH начинается в то время, когда наблюдается вторичное поступление ионов натрия в яйцо и обмен между ионами натрия из морской воды и ионами водорода из яйца в отношении 1:1. В яйце морского ежа обмени-
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. _______________ ОПЛОДОТВОРЕНИЕ ВОЗНИКНОВЕНИЕ НОВОГО ОРГАНИЗМА 65
вается около 75% ионов водорода. Утрата такого количества ионов водорода приводит к повышению pH с 6.8 до 7.2 и к глубоким изменениям физиологии яйца (Shen, Steinhurdt. 1978). Принято считать, что эти изменения обусловлены кальций-зависимой реакцией, однако было обнаружено, что многие из поздних реакций оплодотворения могут вызываться повышением внутриклеточного pH любым способом. Поздние реакции оплодотворения включают активацию синтеза ДНК и белкового синтеза. Резкое повышение белкового синтеза обычно происходит через несколько минут после проникновения спермия и не зависит от синтеза новых мРНК (рис. 2.33). При синтезе новых белков, вероятно, используются мРНК, уже содержащиеся в цитоплазме ооцита. (Более подробно этот вопрос обсуждается в гл. 14.) Подобное скачкообразное увеличение интенсивности белкового синтеза можно вызвать искусственно, помещая неоплодотворенные яйца в раствор, содержащий ионы аммония (Winkler et al., 1980). Эти ионы теряют протоны вне клетки и диффундируют через плазматическую мембрану в виде NH3; проникнув в клетку, они присоединяют протоны и превращаются в NH+4. Отражением происходящей при этом утраты свободных ионов водорода является повышение внутриклеточного pH (рис. 2.34). Таким образом, ионы аммония способны повысить pH цитоплазмы без участия в этом процессе обмена между ионами натрия и водорода При этом они активируют белковый синтез и синтез ДНК. Противоположное действие оказывают агенты, блокирующие повышение pH: одновременно они блокируют поздние события оплодотворения. Если яйца сразу после оплодотворения поместить в морскую воду с низким содержанием ионов натрия или если в морскую воду добавить амилорид (вещество, подавляющее транспорт ионов натрия в клетку), то белковый синтез не активируется, женский и мужской пронуклеусы не перемещаются, деления клетки не происходит (Dube et al.. 1985). 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|