|
|
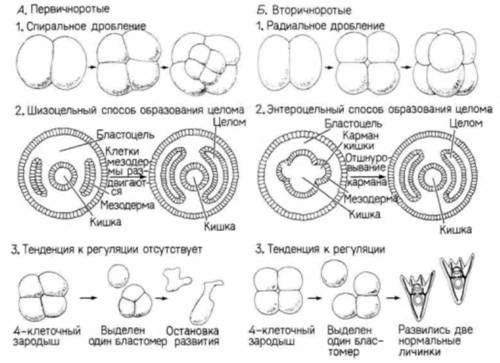
Первичноротые и вторичноротыеДля другой группы многоклеточных, произошедших от колониальных простейших, характерно образование в процессе развития трех зародышевых листков. Некоторые члены этой группы относятся к разделу Radiata, потому что обладают радиальной симметрией подобно трубке или колесу. Radiata включают кишечно-полостных (медузы, кораллы, гидра) и гребневиков. Мезодерма у этих животных находится в зачаточном состоянии: она представлена единичными клетками, разбросанными в студенистом матриксе. Большая часть Metazoa относится, однако, к разделу двусторонне-симметричных животных (Bilateralia) и делится на первично- и вторичноротых. Полагают, что все Bilateralia произошли от примитивных плоских червей. У этих плоских червей впервые появилась истинная мезодерма (хотя она была плотной и в ней не возникала полость тела). Считается, что они были похожи на личинок некоторых современных кишечно-полостных. Различия между двумя основными группами Bilateralia представлены на рис. 1.25. Первичноротые (Protostomia). включающие моллюсков, членистоногих и червей, носят такое название потому, что ротовое отверстие у них возникает из первичного рта (бластопора) или его части. Мезодерма образуется из телобластов. Полость тела образуется путем расхождения первоначально плотного тяжа клеток (схизоцельный способ). Другим большим подразделением двусторонне-симметричных животных являются вторичноротые(Deuterostomia). Они включают хордовых и иглокожих. Может показаться странным включение человека и росомахи в одну систематическую группу с морскими звездами и морскими ежами, однако некоторые эмбриологические особенности делают очевидным их родство. Во-первых, у вторичноротых дефинитивный рот возникает независимо от первичного рта (бластопора) и в другом месте. Мезодерма образуется энтероцельным способом, и имеется вторичная полость тела (целом). Первично- и вторичноротые различаются также по типу дробления. У вторичноротых бластомеры каждого квартета располагаются над соответствую-
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. ВВЕДЕНИЕ В ИЗУЧЕНИЕ РАЗВИТИЯ ЖИВОТНЫХ____ 33
щими бластомерами подлежащего квартета, и анимально-вегетативная ось яйца является у них осью радиальной симметрии. Такой тип дробления называю) радиальным. В отличие от вторичноротых у первичноротых бластомеры вышележащего квартета располагаются между бластомерами нижележащего квартета. Такой тип дробления носит название спирального. Далее, в период дробления, у большинства вторичноротых бластомеры обладают большей способностью к регуляции развития, чем у первичноротых. Если у зародыша морского ежа или человека на 4-клеточной стадии удалить один из бластомеров, то он способен образовать целый организм, а оставшиеся 3/4 зародыша также могут развиваться нормально. Но если такую же операцию проделать на зародыше улитки или червя, то отдельный бластомер и остальная часть зародыша превратятся в двух частичных зародышей; у каждого из них будут отсутствовать те части, которые представлены у партнера. Эволюция организмов зависит от унаследованных изменений в их развитии. Одно из величайших достижений эволюции — возникновение амниотического яйца произошло у вторичноротых. Этот тип яйца, примером которого может служить яйцо курицы (рис. 1.26), впервые появился у рептилий около 30 млн. лет тому назад. Благодаря его возникновению позвоночные в период эмбрионального развития стали независимы от воды. Они получили возможность заселять сушу вдали от водоемов. И если амфибии должны возвращаться к воде для размножения и для того, чтобы их яйца могли развиваться, то яйца амниот имеют собственный запас воды и пиши. Яйца амниот оплодотворяются внутри материнского организма и содержат желток для питания развивающегося зародыша. Кроме того, зародыш амниот снабжен двумя мешками: амнионом, содержащим жидкость, которая его омывает, и аллантоисом. в котором скапливаются конечные продукты обмена. Зародыш вместе с амнионом и аллантоисом заключен в скорлупу, которая проницаема для кислорода, диффундирующего внутрь яйца, но достаточно прочна, чтобы защитить зародыш от неблагоприятных воздействий внешней среды. Сходные эволюционные изменения яйцевых оболочек позволили членистоногим стать первыми сухопутными беспозвоночными. Таким образом, окончательное преодоление границы между водой и сушей явилось результатом изменения самой ранней стадии развития — яйца. Биология развития располагает бесконечным многообразием интереснейших объектов и увлекательных проблем. Задачей автора было отобрать некоторые из них для иллюстрации основных закономерностей развития животных. Выборка эта очень мала. Для того чтобы яснее представить себе состояние дел в эмбриологической науке, можно прибегнуть к такому сравнению: сейчас мы ведем наблюдения только в доступном нам маленьком прудике, тогда как перед нами лежит целый океан неизвестных закономерностей развития. Изучать развитие животных мы начнем с ранних стадий эмбриогенеза: оплодотворения, дробления, гаструляции и установления плана строения тела. Хотя нами была предпринята попытка дать обзор основных вариантов процесса развития во всем животном царстве, легко заметить, что наибольшего внимания удостоились вторичноротые. ЦИТИРОВАННАЯ ЛИТЕРАТУРА
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. ГЛАВА 1
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. Глава 2 Еще, и еще, и еще. Это вечное стремление Вселенной рождать и рождать. Вечно плодородное движение мира. Из мрака выходят двое, они так несхожи, но равны: вечно материя, вечно рост, вечно явление пола. Вечно ткань из различий и тождеств, вечно зарождение жизни УОЛТ УИТМЕН (1866)1
Введение Оплодотворение представляет собой процесс слияния двух половых клеток (гамет)друг с другом, в результате чего возникает новая особь, генетические потенции которой берут начало от обоих родителей. Оплодотворение осуществляет две разные функции: половую (комбинирование генов двух родителей) и репродуктивную (создание нового организма). Первая из этих функций включает передачу генов от родителей потомкам, вторая — инициацию в цитоплазме яйца тех реакций, которые позволяют продолжать развитие. Конкретные особенности оплодотворения очень сильно варьируют у различных видов, но происходящие при этом события обычно сводятся к четырем основным процессам: 1. Контакт спермия с яйцом и их взаимное узнавание. Это этап качественного контроля. Спермий и яйцо должны принадлежать одному и тому же виду. 2. Регуляция проникновения спермия в яйцо. Это этап количественного контроля. Только один спермий должен в конечном счете оплодотворить яйцо. Все остальные спермии должны быть элиминированы. 3. Слияние генетического материала спермия и яйца. 4. Активация метаболизма яйца для его вступления на путь развития. Строение гамет Прежде чем перейти к рассмотрению каждого события, происходящего при оплодотворении, остановимся на строении спермия и яйца, двух типов клеток, которые специализированы для осуществления оплодотворения. Спермий Роль, выполняемая спермием в процессе оплодотворения, была выяснена только в прошлом столетии. Антони ван Левенгук. голландский микроскопист, который был одним из двух ученых, открывших в 1678 г. существование спермиев, первоначально считал их паразитическими животными, живущими, в сперме (отсюда название сперматозоиды, т.е. «животные спермы»). Поначалу он решил, что спермии не имеют никакого отношения к размножению животных, у которых они были обнаружены, но позднее пришел к мысли, что каждый спермий содержит преформированное животное. Левенгук писал (1685), что спермии — это семена (как sperma, так
1 Уолт Уитмен «Листья травы». Гос. изд-во худож. литературы. 1955. с. 49.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 36________________ ГЛАВА 2________________________________________________________________________
и semen означают «семя, зерно») и что самка только предоставляет питательную почву для посева семян. В этом отношении он возвращается к идее порождения, провозглашенной Аристотелем за две тысячи лет до этого. Несмотря на все старания, Левенгуку, к его разочарованию, никак не удавалось обнаружить в сперматозоидах преформированное семя. Николас Хартсекер, открывший спермии одновременно с Левенгуком. нарисовал то, что надеялся найти в спермии: преформированного человечка «гомункулуса» (рис. 2.1). Представление, согласно которому в спермии содержится целый зародыш, никогда не имело много приверженцев, так как оно подразумевает огромное и бессмысленное растрачивание потенциальной жизни. В серии опытов, осуществленных в конце 1700-х годов, Ладзаро Спалланцани показал, что профильтрованная сперма жабы, из которой удалены спермии, не оплодотворяет яиц. Однако он сделал заключение, что действующим началом в оплодотворении является вязкая жидкость, задерживаемая фильтровальной бумагой, а не спермии. Спалланцани тоже полагал, что спермии, содержащиеся в сперме, не что иное, как паразитические животные. И все же проделанный им опыт был первым свидетельством важной роли спермиев при оплодотворении. Улучшение качества линз для микроскопирования и развитие клеточной теории привели к новому пониманию функции спермиев. В 1824 г. Ж. Л. Прево и Ж. Б. Дюма заявили, что спермии — не паразиты, а активные участники оплодотворения. Этими исследователями было установлено неизменное присутствие спермиев у половозрелых самцов и их отсутствие у не достигших зрелости и старых особей. Эти наблюдения в сочетании с известным фактом отсутствия спермиев у стерильных мулов привели к заключению, что «существует тесная зависимость между их присутствием в половых органах и плодовитостью животного». Прево и Дюма утверждали, что на самом деле спермий проникает в яйцо и вносит материальный вклад в формирование следующего поколения. Эти утверждения, как правило, не находили признания, пока в 1840-х годах А. фон Кёлликер не описал образование спермиев из клеток семенника. Он пришел к выводу, что спермии являются сильно измененными клетками, которые образуются в семенниках взрослых самцов. Кёлликер высмеял идею, что наличие спермы может быть нормальным явлением, если она нужна только для того, чтобы поддерживать существование огромного числа паразитов. Несмотря на все это, он считал, что между спермием и яйцом отсутствует какой-либо физический контакт. Он полагал, что спермий побуждает яйцо к развитию подобно тому, как магнит воздействует на железо. И только в 1876 г. Оскар Гертвиг наглядно продемонстрировал проникновение спермия в яйцо и соединение их ядер. Гертвиг разыскивал организм, пригодный для тщательных микроскопических наблюдений, и нашел, что средиземноморский морской еж Toxopneustes lividus прекрасно отвечает его требованиям. Т. lividus был не только обычен в данном районе, и особи этого вида со зрелыми половыми продуктами встречались на протяжении большей части года, но его яйца были доступны в большом числе и были достаточно прозрачны для наблюдений даже при больших увеличениях. После смешивания суспензий спермиев и яиц Гертвиг многократно наблюдал проникновение спермия в яйцо и слияние двух ядер. Он заметил также, что в каждое яйцо проникает только один спермий и что все ядра зародыша являются потомками ядра, возникшего в результате слияния ядер спермия и яйца при оплодотворении. Наблюдения Гертвига были вскоре дополнены Германом Фолем. который проследил в деталях механизм проникновения спермия. Наконец, было признано, что оплодотворение представляет собой слияние спермия и яйца, и соединение гамет
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. _____________ ОПЛОДОТВОРЕНИЕ: ВОЗНИКНОВЕНИЕ НОВОГО ОРГАНИЗМА 37
у морского ежа до сих пор остается наиболее изученным примером оплодотворения. Известно, что каждый спермий содержит гаплоидное ядро, двигательную систему, обеспечивающую перемещение ядра, и мешок, заполненный ферментами, необходимыми для проникновения ядра в яйцо. Большая часть цитоплазмы спермия элиминируется при его созревании, и сохраняются только некоторые органеллы, видоизмененные для выполнения спермием своей функции (рис. 2.2). В период созревания спермия его гаплоидное ядро приобретает обтекаемую форму, а ДНК сильно уплотняется. Впереди от такого конденсированного гаплоидного ядра лежит акросомный пузырек, произошедший от аппарата Гольджи и содержащий ферменты, которые переваривают белки и полисахариды. Таким образом, его можно рассматривать как измененную лизосому. Запас ферментов в акросомном пузырьке служит для проникновения спермия через наружные покровы яйца. V многих видов, например у морских ежей, между ядром и акросомным пузырьком имеется область, содержащая глобулярный актин. На ранних стадиях оплодотворения этот актин используется для образования пальцеобразного выроста. У таких видов молекулы на поверхности акросомного выроста участвуют в узнавании спермием и яйцом друг друга. Акросома и ядро вместе образуют головку спермия. Способ передвижения спермиев различается в зависимости от того, каким образом вид адаптирован
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 38________________ ГЛАВА 2___________________________
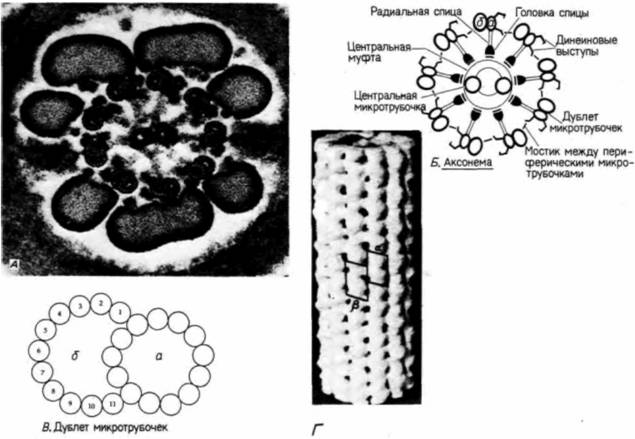
к условиям окружающей среды. У некоторых видов (как, например, у паразитического круглого червя аскариды) спермий перемещается при помощи амебоидного движения ламеллиподий – локальных выростов клеточной мембраны. У большинства видов, однако, спермии способны передвигаться на большие расстояния благодаря биению своих жгутиковЖгутики имеют сложное строение. Главная двигательная структура жгутика называется аксонемойАксонему образуют микротрубочки, отходящие от центриоли, которая находится в основании ядра спермия (рис. 2.2; 2.3, Б). Стержень аксонемы состоит из двух центральных одиночных микротрубочек, окруженных кольцом из девяти двойных микротрубочек (дублетов). В действительности только одна микротрубочка каждого дублета имеет законченное строение и содержит 13 протофиламентов. тогда как вторая имеет С-образное строение и содержит только 11 протофиламентов (рис. 2.3, А— В). Трехмерная модель микротрубочки законченного строения представлена на рис. 2.3, Г. На этом рисунке можно видеть 13 связанных между собой протофиламентов, образованных только димерным белком тубулином. С микротрубочками связан другой белок динеин (рис. 2.3. Б). Этот белок может гидролизовать молекулы аденозинтрифосфата (АТФ) и преобразовывать выделяющуюся при этом химическую энергию в механическую, за счет которой осуществляется движение спермия (Ogawa et. al., 1977). Важная роль белка динеина очевидна на примере мужчин с генетическим синдромом, называемым триадой
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. ОПЛОДОТВОРЕНИЕ ВОЗНИКНОВЕНИЕ НОВОГО ОРГАНИЗМА_____________________________ 39 Картедженера. У таких мужчин динеин отсутствует во всех клетках, обладающих ресничками и жгутиками: соответственно, эти органеллы неподвижны. Мужчины, страдающие таким заболеванием, стерильны (так как спермии неподвижны), подвержены респираторным инфекциям (из-за неподвижности ресничек дыхательных путей), и у них в 50% случаев сердце располагается справа (Afzelius, 1976). Набор микротрубочек типа «9 + 2» с динеиновыми выступами сохранился во всем царстве эукариот. что свидетельствует об исключительной эффективности такой организации для передачи энергии, обеспечивающей движение. Энергия для биения жгутика, благодаря которому осуществляется продвижение спермия, исходит от кольца митохондрий, расположенных в области шейки спермия (рис 2.2). У многих видов (преимущественно у млекопитающих) между митохондриями и аксонемой располагается слой плотных волокон. Этот слой придает хвостовому отделу спермия большую жесткость. Поскольку толщина этого слоя уменьшается по направлению к кончику хвоста, можно думать, что волокна служат для предотвращения слишком резких изгибов головки. Таким образом, для того чтобы перенести материал своего ядра в яйцеклетку, спермий претерпел далеко идущие преобразования. Яйцо Все материалы, необходимые для начальных стадий роста и развития зародыша, должны быть запасены в зрелом яйце.Поэтому, если спермий избавляется от большей части своей цитоплазмы, то развивающееся яйцо не только сохраняет содержащиеся в нем запасные материалы, но и активно увеличивает их количество. Оно либо само синтезирует, либо поглощает белки желтка, который служит источником питания развивающегося зародыша. Так. яйцо птиц представляет собой огромную одиночную клетку, достигшую таких размеров благодаря накоплению желтка. Даже яйца с относительно небольшим содержанием желтка имеют значительные размеры. Объем яйца морского ежа составляет около 2· 105 мкм3 и превосходит объем спермия более чем в 10000 раз. На схематическом рисунке 2.4 показано соотношение их размеров. На этом рисунке изображены также различные структурные элементы зрелого яйца. Если в яйце и спермии содержатся количественно равные гаплоидные ядерные компоненты, то по содержанию цитоплазматических компонентов различия между ними очень велики. Так, цитоплазма яйца представляет собой огромную кладовую запасов, накопленных в период вителлогенеза. Эти запасы включают белки, рибосомы и транспортную
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 40________________ ГЛАВА 2______________________________________________________________________________
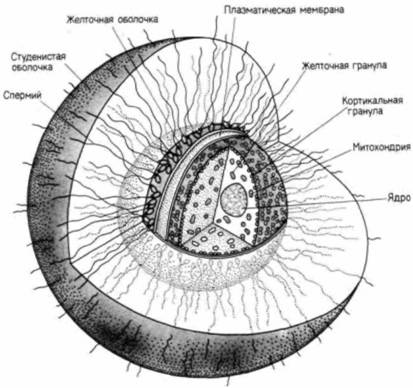
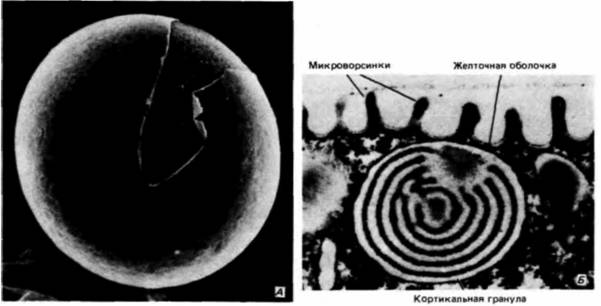
РНК (тРНК), матричную РНК и морфогенетические факторы. 1. Белки. Пройдет еще много времени, прежде чем зародыш окажется способным самостоятельно питаться или получать пишу от матери. Клетки зародыша на ранних стадиях нуждаются в запасенных заранее источниках энергии и аминокислот. У многих видов такими источниками являются накопленные яйцом белки желтка. Многие из желточных белков синтезируются в других органах (печени, жировом теле) и транспортируются к яйцу. 2. Рибосомы и тРНК. Как мы увидим ниже, вскоре после оплодотворения в яйце резко увеличивается синтез белка. Этот синтез осуществляется при участии рибосом и тРНК, которые уже имеются в яйце. Развивающееся яйцо (ооцит) обладает специальным механизмом синтеза рибосом, и в ооцитах некоторых амфибий в течение профазы мейоза образуется до 1012 рибосом. 3. Матричная РНК. У большинства организмов информация для осуществления белкового синтеза, происходящего в раннем развитии, заключена уже в ооците. Полагают, что яйца морского ежа содержат 25—50 тыс. различных типов мРНК. Однако эти мРНК до оплодотворения остаются в неактивном состоянии. 4. Морфогенетические факторы. Эти молекулы, направляющие дифференцировку клеток для образования различных тканей и органов, по-видимому, рассеяны по всему яйцу и распределяются между разными клетками при дроблении (гл. 7 и 8). Внутри этого огромного объема цитоплазмы яйца располагается большое ядро. У некоторых видов животных (например, у морского ежа) ядро к моменту оплодотворения уже гаплоидное. У других видов (в том числе у многих червей и некоторых млекопитающих) ядро яйца в это время еще диплоидное, и спермий проникает в цитоплазму до завершения мейотических делений. Стадии, на которых находится ядро яйца ко времени проникновения спермия, представлены на рис. 2.5. Цитоплазма яйца окружена плазматической мембраной. Во время оплодотворения она регулирует поступление в яйцо некоторых ионов (главным образом Na+) и способна сливаться с плазматической мембраной спермия. К плазматической мембране снаружи примыкает желточная оболочка (рис. 2.6). Образованная гликопротеинами, эта оболочка играет важную роль в обеспечении видовой специфичности прикрепления спермия. У млекопитающих желточная оболочка очень толстая и носит название прозрачной оболочки (zona pellucida). Яйцо млекопитающих окружено также слоем клеток лучистого венца (corona radiata; рис. 2.7). Это фолликулярные клетки, которые доставляли пищу яйцу вплоть до его овуляции. Чтобы оплодотворить яйцо, спермий должен преодолеть также и этот слой 1. Слой цитоплазмы, располагающийся непосредственно под плазматической мембраной яйца морского ежа, называется кортикальным слоем или кортексом. Цитоплазма этого слоя имеет более упорядоченное строение, чем цитоплазма более глубоких слоев, и в ней содержатся высокие концентрации глобулярных молекул актина. При оплодотворении молекулы актина полимеризуются. образуя длинные тяжи, известные под названием микрофиламентов Микрофиламенты необходимы для осуществления клеточного деления: кроме того, они участвуют в образовании выростов, увеличивающих клеточную поверхность.— микроворсинок, которые способствуют проникновению спермия в клетку (рис. 2.6 и 2.18). Кортикальная цитоплазма гораздо более вязкая, чем расположенная глубже основная масса цитоплазмы, и она не смещается при центрифугировании. В кортексе заключены кортикальные гранулы (рис. 2.4; 2.6). Эти мембранные структуры гомологичны акросомному пузырьку спермия, поскольку они содержат протеолитические ферменты и формируются аппаратом Гольджи. Однако если каждый спермий имеет один акросомный пузырек, то каждое яйцо содержит приблизительно 15 000 кортикальных гранул. Кроме того, в кортикальных гранулах помимо пищеварительных ферментов содержатся мукополисахариды и белок гиалинового слоя. Ферменты и мукополисахариды участвуют в защите яйца от проникновения сверхчисленных спермиев после того, как первый спермий придет в контакт с ним; белок гиалинового слоя окружает зародыш на ранних стадиях развития и объединяет бластомеры в период дробления. Яйца многих типов выделяют также студенистое вещество, располагающееся поверх желточной оболочки. Этот студенистый гликопротеиновый слой выполняет различные функции, но одна из главных его функций — это привлечение или активация спер- 1 У некоторых видов млекопитающих во внеклеточных покровах яйца различают три разных образования: I ) прозрачная оболочка (zona pcllucida); 2) яйценосный бугорок (cumulus oophorus) и 3) лучистый венец (corona radiata). В этих случаях лучистым венцом называют те фолликулярные клетки, которые примыкают непосредственно к прозрачной оболочке, а термин яйценосный бугорок применяют к более диффузно распределенным клеткам, расположенным на большем расстоянии от яйца. У многих млекопитающих (например, у хомячка, рис. 2.7. Б) провести границу между яйценосным бугорком и лучистым венцом нелегко; у других (таких, как овца и корова) они, по-видимому, сбрасываются до оплодотворения (см. Talbot, 1985). Клетки лучистого венца и яйценосного бугорка погружены в матрикс, состоящий преимущественно из гиалуроновой кислоты.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. ________________ ОПЛОДОТВОРЕНИЕ: ВОЗНИКНОВЕНИЕ НОВОГО ОРГАНИЗМА_____________________________ 41
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 42________________ ГЛАВА 2_______________
миев. Таким образом, яйцо представляет собой клетку, специализированную для восприятия спермия и инициации развития. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|



 •
•