
|
|
Структура и химический состав клеточного ядра
Ядро неделящейся, интерфазной клетки обычно одно на клетку (хотя встречаются и многоядерные клетки). Ядро состоит из хроматина, ядрышка, кариоплазмы (нуклеоплазмы) и ядерной оболочки, отделяющей его от цитоплазмы (рис. 17). Хроматин При наблюдении живых или фиксированных клеток внутри ядра выявляются зоны плотного вещества, которые хорошо воспринимают разные красители, особенно основные. Благодаря такой способности хорошо окрашиваться этот компонент ядра и получил название «хроматин» (от греч. chroma — цвет, краска). В состав хроматина входит ДНК в комплексе с белком. Такими же свойствами обладают и хромосомы, которые отчетливо видны во время митотического деления клеток. В неделящихся (интерфазных) клетках хроматин, выявляемый в световом микроскопе, может более или менее равномерно заполнять объем ядра или же располагаться отдельными глыбками. Хроматин интерфазных ядер представляет собой хромосомы, которые, однако, теряют в это время свою компактную форму, разрыхляются, деконденсируются. Степень такой деконденсации хромосом может быть различной. Зоны полной деконденсации их участков морфологи называют эухроматином (euchromatinum). При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина, иногда называемого гетерохроматином (heterochromatinum). Степень деконденсации хромосомного материала — хроматина в интерфазе может отражать функциональную нагрузку этой структуры. Чем «диффузнее» распределен хроматин в интерфазном ядре (т.е. чем больше эухроматина), тем интенсивнее в нем синтетические процессы.
Рис. 17. Ультрамикроскопическое строение ядра интерфазной клетки. А — схема; Б — электронная микрофотография участка ядра; 1 — ядерная оболочка (две мембраны, перинуклеарное пространство); 2 — комплекс поры; 3 — конденсированный хроматин; 4 — диффузный хроматин; 5 — ядрышко (гранулярная и фибриллярная части); 6 — межхроматиновые гранулы РНК; 7 — перихроматиновые гранулы; 8 — кариоплазма. Таким образом, хромосомы клеток могут находиться в двух структурно-функциональных состояниях: в активном, рабочем, частично или полностью деконденсированном, когда с их участием в интерфазном ядре происходят процессы транскрипции и редупликации, и в неактивном, в состоянии метаболического покоя при максимальной их конденсированности, когда они выполняют функцию распределения и переноса генетического материала в дочерние клетки. Наблюдения за структурой хроматина с помощью электронного микроскопа показали, что как в препаратах выделенного интерфазного хроматина или выделенных митотических хромосом, так и в составе ядра на ультратонких срезах всегда видны элементарные хромосомные фибриллы толщиной 20—25 нм. В химическом отношении фибриллы хроматина представляют собой сложные комплексы дезоксирибонуклеопротеидов (ДНП), в состав которых входят ДНК и специальные хромосомные белки — гистоновые и негистоновые. В составе хроматина обнаруживается также РНК. Количественные отношения ДНК, белка и РНК составляют 1:1,3:0,2. Обнаружено, что длина индивидуальных линейных молекул ДНК может достигнуть сотен микрометров и даже сантиметров. Среди хромосом человека самая большая первая хромосома содержит ДНК с общей длиной до 7 см. Суммарная длина молекул ДНК во всех хромосомах одной клетки человека составляет около 170 см, что соответствует 6 ×1012 г. В хромосомах существует множество мест независимой репликации ДНК — репликонов. ДНК эукариотических хромосом представляют собой линейные молекулы, состоящие из тандемно (друг за другом) расположенных репликонов разного размера. Средний размер репликона около 30 мкм. В составе генома человека должно встречаться более 50 000 репликонов, участков ДНК, которые синтезируются как независимые единицы. Синтез ДНК как на участках отдельной хромосомы, так и среди разных хромосом идет неодновременно, асинхронно. Так, например, в некоторых хромосомах человека (1, 3, 16) репликация наиболее интенсивно начинается на концах хромосом и заканчивается (при высокой интенсивности включения метки) в центромерном районе (см. ниже). Наиболее поздно репликация заканчивается в хромосомах или в их участках, находящихся в компактном, конденсированном состоянии. Таким примером может являться поздняя репликация генетически инактивированной Х-хромосомы у женщин, формирующей в клеточном ядре компактное тельце полового хроматина.
Белки хроматина составляют 60—70% от его сухой массы. К ним относятся так называемые гистоны и негистоновые белки. Негистоновые белки составляют 20% от количества гистонов. Гистоны — щелочные белки, обогащенные основными аминокислотами (главным образом лизином и аргинином). Очевидна структурная роль гистонов, которые не только обеспечивают специфическую укладку хромосомной ДНК, но и имеют значение в регуляции транскрипции. Гистоны расположены по длине молекулы ДНК не равномерно, а в виде блоков. В один такой блок входят 8 молекул гистонов, образуя так называемую нуклеосому. Размер нуклеосомы около 10 нм. При образовании нуклеосом происходит компактизация, сверхспирализация ДНК, что приводит к укорачиванию длины хромосомной фибриллы примерно в 5 раз. Сама же хромосомная фибрилла имеет вид нитки бус или четок, где каждая бусина — нуклеосома (см. рис. 17). Такие фибриллы толщиной 10 нм дополнительно продольно конденсируются и образуют основную элементарную фибриллу хроматина толщиной 25 нм. Негистоновые белки интерфазных ядер образуют внутри ядра структурную сеть, которая носит название ядерный белковый матрикс, представляющий собой основу, определяющую морфологию и метаболизм ядра. В ядрах, кроме хроматиновых участков и матрикса, встречаются перихроматиновые фибриллы, перихроматиновые и интерхроматиновые гранулы. Они содержат РНК и встречаются практически во всех активных ядрах, представляют собой информационные РНК, связанные с белками, — рибонуклеопротеиды (информосомы). Матрицами для синтеза этих РНК являются разные гены, разбросанные по деконденсированным участкам хромосомных (точнее, хроматиновых) фибрилл. Особый тип матричной ДНК, а именно ДНК для синтеза рибосомной РНК, собран обычно в нескольких компактных участках, входящих в состав ядрышек интерфазных ядер. Ядрышко Практически во всех живых клетках эукариотических организмов в ядре видно одно или несколько обычно округлой формы телец величиной 1—5 мкм, сильно преломляющих свет — это ядрышко, или нуклеола (nucleolus). К общим свойствам ядрышка относится способность хорошо окрашиваться различными красителями, особенно основными. Такая базофилия определяется тем, что ядрышки богаты РНК. Ядрышко — самая плотная структура ядра — является производным хромосомы, одним из ее локусов с наиболее высокой концентрацией и активностью синтеза РНК в интерфазе. Оно не является самостоятельной структурой или органеллой. В настоящее время известно, что ядрышко — это место образования рибосомных РНК (рРНК) и рибосом, на которых происходит синтез полипептидных цепей уже в цитоплазме. Образование ядрышек и их число связаны с активностью и числом определенных участков хромосом — ядрышковых организаторов, которые расположены большей частью в зонах вторичных перетяжек; количество ядрышек в клетках данного типа может изменяться за счет слияния ядрышек или за счет изменения числа хромосом с ядрышковыми организаторами. При исследовании фиксированных клеток вокруг ядрышка всегда выявляется зона конденсированного хроматина, часто отождествляемая с хроматином ядрышкового организатора. Этот околоядрышковый хроматин, по данным электронной микроскопии, представляет собой интегральную часть сложной структуры ядрышка. ДНК ядрышкового организатора представлена множественными (несколько сотен) копиями генов рРНК: на каждом из этих генов синтезируется высокомолекулярный предшественник РНК, который превращается в более короткие молекулы РНК, входящие в состав субъединиц рибосомы. Схему участия ядрышек в синтезе цитоплазматических белков можно представить следующим образом: на ДНК ядрышкового организатора образуется предшественник рРНК, который в зоне ядрышка одевается белком, здесь происходит сборка рибонуклеопротеидных частиц — субъединиц рибосом; субъединицы, выходя из ядрышка в цитоплазму, участвуют в процессе синтеза белка. Ядрышко неоднородно по своему строению: в световом микроскопе можно видеть его тонковолокнистую организацию. В электронном микроскопе выявляются два основных компонента: гранулярный и фибриллярный. Диаметр гранул около 15—20 нм, толщина фибрилл — 6—8 нм. Фибриллярный компонент может быть сосредоточен в виде центральной части ядрышка, а гранулярный — по периферии. Часто гранулярный компонент образует нитчатые структуры — нуклеолонемы толщиной около 0,2 мкм. Фибриллярный компонент ядрышек представляет собой рибонуклеопротеидные тяжи предшественников рибосом, а гранулы — созревающие субъединицы рибосом. В зоне фибрилл можно выявить участки ДНК ядрышковых организаторов. Ультраструктура ядрышек зависит от активности синтеза РНК: при высоком уровне синтеза рРНК в ядрышке выявляется большое число гранул, при прекращении синтеза количество гранул снижается, ядрышки превращаются в плотные фибриллярные тельца базофильной природы. Действие многих веществ (актиномицин, митомицин, ряд канцерогенных углеводородов, циклогексимид, гидрооксимочевина и др.) вызывает в клетках падение интенсивности ряда синтезов и в первую очередь активности ядрышек. При этом возникают изменения в структуре ядрышек: их сжатие, обособление фибриллярных и гранулярных зон, потеря гранулярного компонента, распад всей структуры. Эти изменения отражают степень повреждения ядрышковых структур, связанных главным образом с подавлением синтеза рРНК. Ядерная оболочка Ядерная оболочка (nucleolemma) состоит из внешней ядерной мембраны (m. nuclearis externa) и внутренней мембраны оболочки (m. nuclearis interna), разделенных перинуклеарным пространством, или цистерной ядерной оболочки (cisterna nucleolemmae). Ядерная оболочка содержит ядерные поры (pori nucleares). Мембраны ядерной оболочки в морфологическом отношении не отличаются от остальных внутриклеточных мембран. В общем виде ядерная оболочка может быть представлена как полый двухслойный мешок, отделяющий содержимое ядра от цитоплазмы.
Внешняя мембрана ядерной оболочки, непосредственно контактирующая с цитоплазмой клетки, имеет ряд структурных особенностей, позволяющих отнести ее к собственно мембранной системе эндоплазматической сети: на ней со стороны гиалоплазмы расположены многочисленные рибосомы, а сама внешняя ядерная мембрана может прямо переходить в мембраны эндоплазматической сети. Внутренняя мембрана связана с хромосомным материалом ядра. Наиболее характерными структурами ядерной оболочки являются ядерные поры. Они образуются за счет слияния двух ядерных мембран. Формирующиеся при этом округлые сквозные отверстия поры (annulus pori) имеют диаметр около 80—90 нм. Эти отверстия в ядерной оболочке заполнены сложноорганизованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и этих структур называют комплексом поры (complexus pori) (рис. 18). Такой сложный комплекс поры имеет октагональную симметрию. По границе округлого отверстия в ядерной оболочке располагается три ряда гранул по 8 в каждом: один ряд лежит со стороны ядра, другой — со стороны цитоплазмы, третий расположен между ними в центральной части поры. Размер гранул около 25 нм. От этих гранул отходят фибриллярные отростки. Фибриллы, отходящие от периферических гранул, могут сходиться в центре и создавать как бы перегородку, диафрагму поперек поры (diaphragma pori). Размеры пор у данной клетки обычно стабильны, так же как относительно стабилен размер ядерных пор клеток разных организмов.
Число ядерных пор зависит от метаболической активности клеток: чем интенсивнее синтетические процессы в клетках, тем больше пор на единицу поверхности клеточного ядра. Так, у эритробластов (клеток-предшественников ядерных эритроцитов) низших позвоночных животных во время интенсивного синтеза и накопления гемоглобина обнаруживается в ядре около 30 ядерных пор на 1 мкм2. После того как эти процессы заканчиваются, в ядрах зрелых клеток — эритроцитов прекращается синтез ДНК и РНК и количество пор снижается до 5 на 1 мкм2. В ядерных оболочках полностью зрелых сперматозоидов поры не обнаруживаются. Из многочисленных свойств и функциональных нагрузок ядерной оболочки следует подчеркнуть ее роль как барьера, отделяющего содержимое ядра от цитоплазмы, ограничивающего свободный доступ в ядро крупных агрегатов биополимеров, регулирующего транспорт макромолекул между ядром и цитоплазмой. Одной из важных функций ядерной оболочки следует считать ее участие в создании внутриядерного порядка — в фиксации хромосомного материала в трехмерном пространстве ядра. В интерфазе часть хроматина структурно связана с внутренней ядерной мембраной. Описаны случаи примембранной локализации центромерных и теломерных участков интерфазных хромосом. Некоторые термины: · болезни хромосомные -- болезни, возникающие вследствие изменения числа или нарушения структуры хромосом в гаметах родителей или на ранних стадиях дробления зиготы; · мутация (-и) (лат. mutatio изменение, перемена) -- всеобщее свойство живых организмов, лежащее в основе эволюции и селекции всех форм жизни и заключающееся во внезапном изменении генетической информации; · мутационное равновесие -- равновесие между частотой возникновения и скоростью элиминации мутаций в популяции; · половой хроматин женский (син.: тельце Барра , X-хроматин) -- плотно окрашиваемый крупный участок хромосомы треугольной, круглой или палочковидной формы, расположенный на периферии ядра соматической клетки женского организма; представляет собой конденсированную X-хромосому; · хроматин половой мужской (син. Y-хроматин) -- крупная глыбка хромосомы в ядре соматической клетки мужского организма, ярко светящаяся при окраске флюорохромами -- производными акридинового оранжевого, напр. акрихином; представляет собой конденсированный участок длинного плеча Y-хромосомы;

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|

 Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде плотных хромосом. В этот период хромосомы не выполняют никаких синтетических функций, в них не происходит включения предшественников ДНК и РНК.
Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде плотных хромосом. В этот период хромосомы не выполняют никаких синтетических функций, в них не происходит включения предшественников ДНК и РНК.
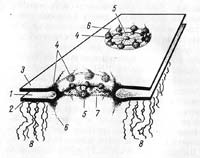 Рис. 18. Строение комплекса поры (схема). 1 — перинуклеарное пространство; 2 — внутренняя ядерная мембрана; 3 — наружная ядерная мембрана; 4 — периферические гранулы; 5 — центральная гранула; 6 — фибриллы, отходящие от гранул; 7 диафрагма поры; 8 — фибриллы хроматина.
Рис. 18. Строение комплекса поры (схема). 1 — перинуклеарное пространство; 2 — внутренняя ядерная мембрана; 3 — наружная ядерная мембрана; 4 — периферические гранулы; 5 — центральная гранула; 6 — фибриллы, отходящие от гранул; 7 диафрагма поры; 8 — фибриллы хроматина.