
|
|
Межклеточные соединения (контакты)
2 — пальцевидное соединение; 3 — десмосома; 4 — плотное соединение; 5 — щелевидное соединение (нексус).
Плазмолемма многоклеточных животных организмов принимает активное участие в образовании специальных структур — межклеточных соединений (junctiones intercellulares), обеспечивающих межклеточные взаимодействия. Различают несколько типов таких структур (рис. 7). Простое межклеточное соединение, (junctio intercellularis simplex) — сближение плазмолемм соседних клеток на расстояние 15—20 нм. При этом происходит взаимодействие слоев гликокаликса соседних клеток. Разновидностью простого соединения является "пальцевидное", или соединение по типу замка. Плотное соединение (запирающая зона) (zonula occludens) — зона, где слои двух плазмолемм максимально сближены, здесь происходит как бы слияние участков плазмолемм двух соседних клеток. Роль плотного замыкающего соединения заключается в механическом соединении клеток друг с другом. Эта область непроницаема для макромолекул и ионов и, следовательно, она запирает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды. Часто встречается, особенно в эпителии, особый тип соединения — пятно сцепления, или десмосома (desmosoma). Эта структура представляет собой небольшую площадку, иногда имеющую слоистый вид, диаметром до 0,5 мкм, где между мембранами располагается зона с высокой электронной плотностью. К плазмолемме в зоне десмосомы со стороны цитоплазмы прилегает участок электронноплотного вещества, так что внутренний слой мембраны кажется утолщенным. Под этим утолщением находится область тонких фибрилл, которые могут быть погружены в относительно плотный матрикс. Функциональная роль десмосом заключается главным образом в механической связи между клетками. Щелевидное соединение, или нексус (nexus), представляет собой область протяженностью 0,5—3 мкм, где плазмолеммы разделены промежутком в 2—3 нм. Со стороны цитоплазмы никаких специальных примембранных структур в данной области не обнаруживается, но в структуре плазмолемм соседних клеток друг против друга располагаются специальные белковые комплексы (коннексоны), которые образуют как бы каналы из одной клетки в другую. Этот тип соединения встречается во всех группах тканей. Функциональная роль щелевидного соединения заключается в переносе ионов и мелких молекул от клетки к клетке. Так, в сердечной мышце возбуждение, в основе которого лежит процесс изменения ионной проницаемости, передается от клетки к клетке через нексус. Синаптические соединения, или синапсы (synapsis). Этот тип соединений характерен для нервной ткани и встречается в специализированных участках контакта как между двумя нейронами, так и между нейроном и каким-либо иным элементом, входящим в состав рецептора или эффектора (например, нервно-мышечные, нервно-эпителиальные синапсы). Синапсы — участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому. Строение синапса также рассматривается в теме нервные окончания. Некоторые термины: · органоид (гр. organon орудие, орган; + гр. -eides подобный; син. органелла) -- постоянная цитоплазматическая структура животной или растительной клетки, выполняющая определенную функцию (напр., митохондрия, рибосома, клеточный центр); · фагоцитоз (phagocytosis; фагоцит + -оз) -- процесс активного захватывания и поглощения микроорганизмов, разрушенных клеток и инородных частиц одноклеточными организмами или особыми клетками (фагоцитами) многоклеточных организмов; · пиноцитоз (pinocytosis; греч. pineo- пить, поглощать + гист. cytus клетка + -оз) -- активное поглощение клеткой жидкости из окружающей среды с формированием в цитоплазме пузырьков, содержащих жидкость; · экзоцитоз (экзо- + гист. cytus клетка + -оз) -- процесс выделения клеткой вещества в виде секреторных гранул или вакуолей; · секреция (secretio; лат. "отделение", "выделение") -- процесс выработки клеткой секрета и выделения его на поверхность эпителия или во внутреннюю среду организма; · экскреция (лат. excerno, excretum, отделять, выделять; от экс- + cerno - различать, просеивать; син. выделение) -- совокупность физиологических процессов, направленных на освобождение организма от конечных продуктов обмена, чужеродных веществ, а также избытка воды, минеральных и органических веществ, поступивших с пищей или образовавшихся в организме в процессе метаболизма; · экскрет (лат. excerno, excretum отделять, выделять) -- конечный продукт обмена веществ, выделяемый организмом;
Часть третья: органеллы; мембранные органеллы цитоплазмы. Органеллы цитоплазмы Органеллы — постоянно присутствующие и обязательные для всех клеток микроструктуры, выполняющие жизненно важные функции. Классификация органелл. Различают мембранные и немембраные органеллы. К мембранным органеллам относятся митохондрии, эндоплазматическая сеть, аппарат Гольджи, лизосомы. Немембранные органеллы: свободные рибосомы и полисомы, микротрубочки, центриоли и филаменты (микрофиламенты, промежуточные филаменты). Во многих клетках органеллы могут принимать участие в образовании особых структур, характерных для специализированных клеток. Так, реснички и жгутики образуются за счет центриолей и плазматической мембраны, микроворсинки — это выросты плазматической мембраны с гиалоплазмой и микрофиламентами, акросома спермиев — это производное элементов аппарата Гольджи, «эллипсоид» зрительных клеток — скопления митохондрий и пр. Мембранные органеллы Мембранные органеллы представляют собой одиночные или связанные друг с другом отсеки цитоплазмы, отграниченные мембраной от окружающей их гиалоплазмы, имеющие свое собственное содержимое, отличное по составу, свойствам и функциям от других частей клетки, т.е. это замкнутые, закрытые объемные зоны — компартменты. В гиалоплазме мембранные органеллы распределены закономерно. Эндоплазматическая сеть, различные вакуоли, возникающие из нее, составляют вакуолярную систему цитоплазмы, систему синтеза и внутриклеточного транспорта веществ. Кроме того, в ее состав входят комплекс Гольджи, лизосомы, аутолизосомы и пероксисомы. Для всех элементов вакуолярной системы характерно наличие одной ограничивающей мембраны. Эндоплазматическая сеть Эндоплазматическая сеть (ЭПС) была открыта К.Р. Портером в 1945 г. Эта мембранная органелла общего назначения представляет собой совокупность вакуолей, плоских мембранных мешков или трубчатых образований, создающих как бы мембранную сеть внутри цитоплазмы. Различают два типа — гранулярную (шероховатую) и гладкую эндоплазматическую сеть.
А — схема; Б — электронная микрофотография участка среза печеночной клетки. 1 — рибосомы; 2 — пластинки; 3 — внутренние полости цистерн; 4 — отщепляющиеся мембранные пузырьки, лишенные рибосом. Гранулярная эндоплазматическая сеть (reticulum endoplasmaticum granulosum) представлена замкнутыми мембранами, которые образуют уплощенные мешки, цистерны, трубочки. Ширина полостей цистерн значительно варьирует в зависимости от функциональной активности клетки. Наименьшая ширина их — около 20 нм, но они могут достигать диаметра в несколько микрометров. Отличительной чертой мембран гранулярной ЭПС является то, что они со стороны гиалоплазмы покрыты рибосомами (рис. 8). Гранулярная эндоплазматическая сеть бывает представлена редкими разрозненными цистернами или их локальными скоплениями. Первый тип гранулярной эндоплазматической сети, характерен для малоспециализированных ктеток или для клеток с низкой метаболической активностью. Скопления эндоплазматической сети являются принадлежностью клеток, активно синтезирующих секреторные белки. Так, в клетках печени и некоторых нервных клетках гранулярная эндоплазматическая сеть собрана в отдельные зоны. В клетках поджелудочной железы гранулярная эндоплазматическая сеть в виде плотно упакованных друг около друга мембранных цистерн занимает базальную и околоядерную зоны клетки. Рибосомы, связанные с мембранами эндоплазматической сети, участвуют в синтезе белков, выводимых из данной клетки («экспортируемые» белки). Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков — ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения. Белки, накапливающиеся в полостях эндоплазматической сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они часто модифицируются и входят в состав либо лизосом, либо секреторных гранул, содержимое которых остается изолированным от гиалоплазмы мембраной. В ряде случаев внутри самих канальцев или вакуолей гранулярной эндоплазматической сети может происходить модификация белков, например связывание их с сахарами (глюкозилирование), или конденсация синтезированных белков с образованием крупных агрегатов — секреторных гранул. В гранулярной эндоплазматической сети происходит синтез мембранных интегральных белков (см. часть 2), которые встраиваются в толщу мембраны. Итак, роль гранулярной эндоплазматической сети заключается в синтезе на ее рибосомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, в химической модификации таких белков и в их локальной конденсации, а также в синтезе структурных компонентов клеточных мембран. Агранулярная (гладкая) эндоплазматическая сеть (reticulum endoplasmaticum nongranulosum) также представлена мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев гладкой эндоплазматической сети обычно около 50—100 нм. Гладкая эндоплазматическая сеть возникает и развивается за счет гранулярной эндоплазматической сети (при освобождении ее от рибосом). Деятельность гладкой эндоплазматической сети связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая эндоплазматическая сеть участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих такие категории липидов, как стероиды, например, в клетках коркового вещества надпочечников, в сустентоцитах семенников. Тесная топографическая связь гладкой эндоплазматической сети с отложениями гликогена (запасной внутриклеточный полисахарид животных) в гиалоплазме различных клеток (клетки печени, мышечные волокна) указывает на ее возможное участие в метаболизме углеводов. В поперечнополосатых мышечных волокнах гладкая эндоплазматическая сеть способна депонировать ионы кальция, необходимые для функции мышечной ткани.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
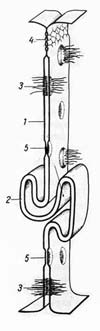 Рис. 7. Межклеточные соединения (схема). 1 — простое соединение;
Рис. 7. Межклеточные соединения (схема). 1 — простое соединение;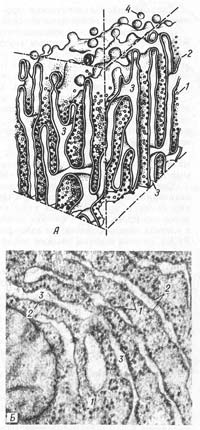 Рис. 8. Строение гранулярной эндоплазматической сети.
Рис. 8. Строение гранулярной эндоплазматической сети. Очень важна роль гладкой эндоплазматической сети в дезактивации различных вредных для организма веществ за счет их окисления с помощью ряда специальных ферментов. Особенно четко она проявляется в клетках печени. Так, при ряде отравлений в клетках печени появляются ацидофильные зоны (не содержащие РНК), сплошь занятые гладким эндоплазматическим ретикулумом.
Очень важна роль гладкой эндоплазматической сети в дезактивации различных вредных для организма веществ за счет их окисления с помощью ряда специальных ферментов. Особенно четко она проявляется в клетках печени. Так, при ряде отравлений в клетках печени появляются ацидофильные зоны (не содержащие РНК), сплошь занятые гладким эндоплазматическим ретикулумом.