
|
|
Микротрубочки интерфазных клетокПрактически во всех эукариотических клетках в гиалоплазме можно видеть длинные неветвящиеся микротрубочки. В больших количествах они обнаруживаются в цитоплазматических отростках нервных клеток, фибробластов и других изменяющих свою форму клеток. Они могут быть выделены сами или можно выделить образующие их белки: это те же тубулины со всеми их свойствами. Главное функциональное значение таких микротрубочек цитоплазмы заключается в создании эластичного, но одновременно устойчивого внутриклеточного каркаса (цитоскелета), необходимого для поддержания формы клетки. Действие яда колхицина, вызывающего деполимеризацию тубулинов, сильно меняет форму клеток. Так, если отростчатую и плоскую клетку в культуре фибробластов обработать колхицином, то она теряет полярность и сжимается. Точно так же ведут себя другие клетки: колхицин прекращает рост клеток хрусталика, отростков нервных клеток, образование мышечных трубок и др. Так как при этом не исчезают элементарные формы движения, присущего клеткам, в частности пиноцитоз, ундулирующие движения мембран, образование мелких псевдоподий, вероятнее всего, роль микротрубочек заключается в образовании каркаса для поддержания формы клеточного тела, для стабилизации и укрепления клеточных выростов. Создавая внутриклеточный скелет, микротрубочки могут быть факторами ориентированного движения клетки в целом и ее внутриклеточных компонентов, задавать своим расположением векторы для направленных потоков разных веществ и для перемещения крупных структур. Разрушение микротрубочек, например колхицином, нарушает транспорт веществ в аксонах нервных клеток, приводит к блокаде секреции и другим нарушениям транспорта веществ. С цитоплазматическими микротрубочками связаны специальные белки, участвующие в механическом переносе отдельных внутриклеточных компонентов: микровакуолей, рибосом, митохондрий, других органелл. В неделящейся (интерфазной) клетке система микротрубочек развивается в связи с особой клеточной органеллой — центриолью, которая является местом, где происходит начальная полимеризация тубулинов и рост микротрубочек цитоскелета. Центриоли Этот термин был предложен Т. Бовери в 1895 г. для обозначения очень мелких телец, размер которых находится на границе разрешающей способности светового микроскопа. В некоторых объектах удавалось видеть, что мелкие плотные тельца — центриоли (centriolum), обычно расположенные в паре — т.н. диплосома (diplosoma), окружены зоной более светлой цитоплазмы, от которой отходят радиально тонкие фибриллы (т.н. центросфера). Совокупность центриолей и центросферы называют клеточным центром (cytocentrum). Эти органеллы в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся (интерфазных) клетках центриоли часто определяют полярность клеток, например, эпителия, и располагаются вблизи комплекса Гольджи.
А — схема; Б — электронная микрофотография; 1 — активная материнская центриоль, окруженная тонкофибриллярным матриксом, от которого отходят микротрубочки полярной лучистости (2); 3 — неактивная дочерняя центриоль.
Тонкое строение центриолей удалось изучить только с помощью электронного микроскопа. Основой строения центриолей являются расположенные по окружности 9 триплетов микротрубочек (triplomicrotubuli), образующих таким образом полый цилиндр. Его ширина около 0,2 мкм, а длина — 0,3—0,5 мкм (хотя встречаются центриоли, достигающие в длину нескольких микрометров) (рис. 14). Кроме микротрубочек в состав центриоли входят дополнительные структуры — «ручки», соединяющие триплеты. Соединительные «ручки» построены из белка динеина, обладающего АТФ-азной активностью и обеспечивающего движение микротрубочек друг относительно друга. Системы микротрубочек центриоли можно описать формулой: (9 x 3) + 0, подчеркивая отсутствие микротрубочек в ее центральной части. Обычно в интерфазных клетках всегда присутствуют две центриоли, располагающиеся рядом друг с другом, образуя диплосому. В диплосоме центриоли располагаются под прямым углом по отношению друг к другу. Из двух центриолей различают материнскую и дочернюю. Обе центриоли сближены и расположены так, что конец дочерней центриоли направлен к поверхности материнской центриоли.
При подготовке клеток к митотическому делению происходит удвоение центриолей. Этот процесс у различных объектов происходит в разное время — в течение синтеза ядерной ДНК или после него. Он заключается в том, что две центриоли в диплосоме расходятся и около каждой из них возникает заново по одной новой дочерней, так что в клетке перед делением обнаруживаются две диплосомы, т.е. четыре попарно связанные центриоли. Этот способ увеличения числа центриолей был назван дупликацией. Важно отметить, что увеличение числа центриолей не связано с их делением, почкованием или фрагментацией, а происходит путем образования зачатка, процентриоли, вблизи и перпендикулярно к исходной центриоли. Полагают, что центриоли участвуют в индукции полимеризации тубулина при образовании микротрубочек. Так, в интерфазе именно в связи с центриолью происходит рост микротрубочек клеточного каркаса. Перед митозом центриоль является одним из центров полимеризации микротрубочек веретена клеточного деления. Центриоль — центр роста микротрубочек аксонемы ресничек или жгутиков. Наконец, она сама индуцирует полимеризацию тубулинов новой процентриоли, возникающей при ее дупликации. Реснички и жгутики Это специальные органеллы движения, встречающиеся в некоторых клетках различных организмов. В световом микроскопе эти структуры выглядят как тонкие выросты клетки. В основании ресничек (cilium) и жгутика flagellum) в цитоплазме видны хорошо красящиеся мелкие гранулы — базальные тельца (corpusculum basale). Длина ресничек 5—10 мкм, а длина жгутиков может достигать 150 мкм (см. рис. 43, В). Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 200 нм. Этот вырост от основания до самой его верхушки покрыт плазматической мембраной. Внутри выроста расположена аксонема («осевая нить») — сложная структура, состоящая в основном из микротрубочек. Проксимальная часть реснички (базальное тело) погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 150 нм). Базальное тельце по своей структуре очень сходно с центриолью. Оно также состоит из 9 триплетов микротрубочек, имеет динеиновые «ручки». Часто в основании реснички лежит пара базальных телец, располагающихся под прямым углом друг к другу подобно диплосоме. Аксонема (filamentum axiale) в своем составе имеет в отличие от базального тельца или центриоли 9 дублетов микротрубочек с динеиновыми «ручками», образующих стенку цилиндра аксонемы. Кроме периферических дублетов микротрубочек, в центре аксонемы располагается пара центральных микротрубочек. В целом систему микротрубочек реснички описывают как (9 x 2) + 2 в отличие от (9 x 3) + 0 системы центриолей и базальных телец. Базальное тельце и аксонема структурно связаны друг с другом и составляют единое целое: две микротрубочки триплетов базального тельца являются микротрубочками дублетов аксонемы. Свободные клетки, имеющие реснички и жгутики, обладают способностью двигаться, а неподвижные клетки движением ресничек могут перемещать жидкость и частицы. Траектория движения ресничек очень разнообразна. В различных клетках это движение может быть маятникообразным, крючкообразным, воронкообразным или волнообразным. Основной белок ресничек — тубулин — не способен к сокращению, укорочению. Вероятным кандидатом на роль сократимого белка считается белок «ручек» — динеин, так как он обладает АТФ-азной активностью. В последние годы для объяснения способа движения ресничек и жгутиков используется гипотеза «скользящих нитей». Известно, что сокращение мышечных волокон происходит за счет встречного скольжения фибрилл двух мышечных белков: миозина и актина; при этом также не происходит собственно укорачивания или сокращения отдельных мышечных белковых фибрилл. Предполагается, что незначительные смещения дублетов микротрубочек друг относительно друга могут вызвать изгиб всей реснички, а если такое локальное смещение будет происходить вдоль жгутика, то может возникнуть волнообразное его движение. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
 Рис. 14. Строение клеточного центра в полюсе митотического веретена клетки.
Рис. 14. Строение клеточного центра в полюсе митотического веретена клетки.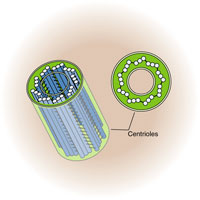 Вокруг каждой центриоли расположен бесструктурный, или тонковолокнистый, матрикс. Часто можно обнаружить несколько дополнительных структур, связанных с центриолями: спутники (сателлиты), фокусы схождения микротрубочек, дополнительные микротрубочки, образующие особую зону, центросферу вокруг центриоли.
Вокруг каждой центриоли расположен бесструктурный, или тонковолокнистый, матрикс. Часто можно обнаружить несколько дополнительных структур, связанных с центриолями: спутники (сателлиты), фокусы схождения микротрубочек, дополнительные микротрубочки, образующие особую зону, центросферу вокруг центриоли.