
|
|
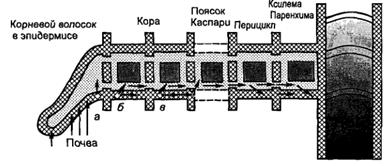
Ближний и дальний транспорт ионов в растенияхВ зависимости от уровня организации процесса различают три типа транспорта веществ в растении: внутриклеточный, ближний (внутри органа) и дальний (между органами). Внутриклеточный транспорт. Напомним, что ионы из почвенного раствора сначала попадают в свободное пространство клеточной стенки. Затем часть их транспортируется через плазмалемму в протопласт с помощью транспортных белков. Транспортные белки действуют на очень небольшом расстоянии – 80 нм. Передвижение веществ внутри клетки осуществляется благодаря круговому движению цитоплазмы (циклозису) и направленной поперек этого движения диффузии. У высших растений движение цитоплазмы происходит с помощью микротрубочек и микрофиламентов. Скорость движения цитоплазмы, достигающая 0,2–0,6 мм/мин, зависит от физиологического состояния клетки и прежде всего от количества АТФ. Вещества, вызывающие разрушение микрофиламентов и разборку микротрубочек, влияют и на внутриклеточный транспорт ионов. Во внутриклеточном транспорте веществ принимают участие также каналы эндоплазматического ретикулума и везикулы Гольджи. Ближний транспорт в корне. Поглощенные клетками эпиблемы корня ионы и продукты их ассимиляции могут двигаться к сосудам центрального цилиндра двумя путями: по симпласту и апопласту. Корневой волосок – это главный вход в симпласт. Из корневого волоска или других клеток эпиблемы поглощенный ион переходит из одной клетки коры в другие по плазмодесмам. Число плазмодесм, связывающих самые мелкие из этих клеток, достигает 20 000–30 000, а иногда даже – 100 000. Внутри симпласта ионы и их органические производные передвигаются от клетки к клетке по плазмодесмам без преодоления мембран со скоростью 1,5–2,0 см/ч. Войдя в протопласт, ион может затем попасть внутрь эндоплазматического ретикулума. Так как в состав каждой плазмодесмы входит его канал, то попавший в него ион может транспортироваться по симпласту из клетки в клетку, не выходя в цитозоль.
Какую роль играют вакуоли в транспорте веществ по симпласту? Ионы не идут через вакуоли, но вакуоли и проводящие ткани корня конкурируют между собой за поглощаемые вещества. Результат конкуренции определяется степенью насыщенности тканей растения солями. У высоко-солевых растений, т.е. у растений, вакуоли которых насыщены ионами, основная часть вновь поглощаемых ионов передвигается в сосуды, минуя встречаю-щиеся вакуоли. У низкосолевых растений, т.е. выращенных на разбавленных питательных растворах, большая часть поглощаемых ионов накапливается в вакуолях корневых клеток, что надолго исключает их из прямого транспорта в сосуды. На продолжительность нахождения иона в клеточном соке влияет его концентрация в почвенном или питательном растворе. Если во внешней среде содержится оптимальное количество иона, то вакуоль не оказывает влияния на его транспорт по симпласту. Если во внешней среде недостаток иона или его нет совсем, то он поступает из вакуоли в цитозоль и потом в симпластический путь. Если во внешней среде избыток иона, то происходит его накопление в вакуоли. Таким образом, вакуоли поддерживают концентрацию данного иона в симпласте постоянной. Поглощение ионов вакуолями снижает их концентрацию в симпласте и обеспечивает создание градиента концентрации, необходимого для транспорта их по симпласту. Таким образом, корень способен выполнять распределительную функцию, направляя в побеги дефицитные элементы питания и задерживая в вакуолях избыточные или вредные. Это очень важно, так как известно, что по мере старения клеток корня их поглотительная способность падает, однако корень продолжает Если концентрация соли в наружном растворе невелика, то большинство ионов, поступающих в корень, попадает к транспортным белкам и далее, пройдя через плазмалемму, – в симпласт. При увеличении наружной концентрации соли транспортные белки плазмалеммы оказываются в какой-то момент «занятыми». При этом часть ионов с током воды поступает в апопласт. Основным путем апопластного транспорта являются рыхлые первичные клеточные стенки. Однако и в апопласте свобода передвижения ограничена и объясняется тем, что карбоксильные группы клеточной стенки несут отрицательный заряд, поэтому катионы легко адсорбируются на поверхности и могут далее мигрировать по ней в глубь корня, анионы же отталкиваются одноименными зарядами клеточной стенки и поэтому проникают в апопласт труднее. В результате неодинакового проникновения катионов и анионов через клеточные стенки внутри корня создается избыток катионов, а следовательно, положительный потенциал, препятствующий их дальнейшему проникновению. Между апопластом и симпластом может идти обмен ионами. Он происходит в области пор, где вторичные стенки отсутствуют. Вода с растворенными веществами, находящаяся в апопласте, как бы омывает все клетки коры корня. Если ион, двигаясь по апопласту, встретит свободный белок-переносчик в плазмалемме, то он может пройти в протопласт клетки коры. Следовательно, длина пути, пройденного ионом по апопласту, определяется числом и скоростью работы транспортных белков. Таким образом, ион может попасть в симпласт не только через корневой волосок, но и через любую клетку коры в глубине корня. В протопласте ион включается в обмен веществ или продолжает свое движение уже по симпласту. Передвигаясь по апопласту, вещества, как и вода, встречают меньшее сопротивление, чем при движении через плазмалемму. Ионы, как и молекулы воды, по апопласту могут двигаться до эндодермы. В эндодерме происходит обязательный переход ионов с апопластного на симпластный путь. Для этого вещества должны пройти через плазмалемму клеток эндодермы, а это в основном активный процесс. Эта необходимость переключения транспорта с апопластного на симпластный путь имеет большое значение, так Растворенное вещество, оказавшееся в протопласте эндодермальной клетки и не включившееся здесь в обмен веществ, либо перемещается дальше из клетки в клетку по симпласту, либо вновь проходит через плазмалемму в какой-нибудь точке с внутренней стороны от пояска Каспари и после этого двигается опять по апопласту. Основным механизмом передвижения по апопласту тоже считают диффузию и массовый ток. Следовательно, скорость транспорта веществ по апопласту зависит от скорости транспирации. Доказательством того, что вещества передвигаются и по апопласту, и по симпласту, являются следующие факторы: I) существование поясков Каспари, прерывающих движение по апопласту; 2) в процессе движения ионов происходят их превращения, а это возможно лишь в симпласте; 3) ионы могут перемещаться быстро и на большие расстояния в отсутствие массового тока, причем вода и вещества передвигаются с разной скоростью; 4) о транспорте ионов по симпласту свидетельствует и зависимость этого процесса от температуры, снабжения ассимилятами, интенсивности света, присутствия в среде ингибиторов метаболизма. В процессе транспорта веществ в корне, каким бы ни был их основной путь – по симпласту или по апопласту, – они, по крайней мере, дважды должны пройти через плазмалемму: на входе в симпласт и на выходе из него. В более сложных случаях движение через симпласт может быть связано с прохождением через мембраны эндоплазматической сети или каких-нибудь других органелл. Доля участия каждого пути в транспорте веществ очень сильно зависит от физиологического состояния растения, интенсивности транспирации и концентрации наружного раствора. Транспорт по апопласту зависит прежде всего от движения воды через корень, поэтому этот путь называют водозависимым. Транспорт по апопласту может играть существенную роль в крайних условиях: повышенная транспирация при высокой температуре, высокая концентрация наружного раствора при внесении удобрений, высокая насыщенность тканей солями. Симпластический транспорт преобладает в оптимальных условиях. Симпластный путь является главным для многих ионов, при этом некоторые из них, особенно азот и фосфор, во время транспорта включаются в химические соединения. В результате эти элементы продолжают свой путь к проводящим тканям корня уже не в свободной форме, а в составе органических веществ. Движение ионов к сосудам происходит довольно быстро. Конечным этапом ближнего транспорта в корне является выделение ионов и продуктов их первичной ассимиляции в сосуды. До сих пор не решен вопрос о механизме поступления ионов в мертвые сосуды ксилемы. Общий характер ближнего транспорта ионов в корне можно представить так: анионы, а в определенных условиях и ионы калия передвигаются в сосуды активно, с помощью транспортных белков, а катионы натрия, кальция, магния пассивно диффундируют вдоль электрохимического градиента, который возникает в результате действия этих насосов. Поступление ионов и молекул в сосуды ксилемы может происходить не только пассивно, но и активно, с помощью транспортных белков. Поступление веществ в сосуды зависит не только от скорости поглощения ионов, но и от интенсивности дыхания и возраста корня. С возрастом в составе пасоки увеличивается доля элементов, запасенных ранее в вакуолях. Корень регулирует равномерность поступления в надземные органы элементов минерального питания. Минеральные ионы и продукты их первичной ассимиляции поступают в сосуды, за ними по законам осмоса входит вода, и возникает корневое давление. Транспирация и корневое давление способствуют массовому току элементов минерального питания по сосудам в побеги. Ионы, попавшие в проводящие ткани побега, вновь поглощаются клетками стебля или листьев. Ближний транспорт в листе. Крупные проводящие пучки, переходящие из стебля в лист, либо разветвляются в нем на все более мелкие пучки второго, третьего и других порядков (у двудольных), либо проходят вдоль всей листовой пластинки в виде параллельных линий, связанных тончайшими боковыми жилками, которые делят основную часть листа на отдельные мелкие участки (у однодольных). Центр любого такого участка отделен от ближайшего окончания проводящей системы всего лишь несколькими клетками. Проводящие пучки окружены обкладкой, которая отделяет их от мезофилла. Обкладка представляет собой один слой видоизмененных паренхимных клеток. Она, особенно у однодольных, напоминает эндодерму корня: у представителей некоторых семейств в радиальных стенках клеток обкладки имеется структура, похожая на поясок Каспари. После того как ксилемный сок (пасока) достигнет мельчайших ответвлений проводящей системы, вода и растворенные в ней вещества поступают в стенки клеток хлоренхимы. Концентрация его определяется скоростью транспирации. При интенсивной транспирации раствор, достигающий плазмалеммы клеток мезофилла, концентрированнее того, который омывает клетки коры корня. При слабой транспирации этот раствор может иметь ту же концентрацию, что и пасока. Из листовых жилок вещества поступают в свободное пространство клеток мезофилла, а потом в симпласт. Другими словами, вещества должны пройти путь, обратный тому, по которому они передвигаются, поступая в проводящую ткань корня. Поступление ионов в симпласт и в свободное пространство листа должно регулироваться скоростью их прохождения через плазмалемму живых клеток. Дальний транспорт поглощенных из почвы ионов – это восходящий ток – он идет не только по сосудам, но и по ситовидным трубкам. В естественных условиях движение веществ по коре или по древесине также зависит от содержания в почве солей. Если концентрация иона в почвенном растворе высокая, то он будет передвигаться по ситовидным трубкам и по сосудам, но если низкая, – только по ситовидным трубкам. Чем меньше в почве данного иона, тем большая его часть транспортируется по ситовидным трубкам. Увеличение концентрации солей в почвенном растворе приводит к увеличению концентрации ксилемного сока, концентрация же веществ, двигающихся по ситовидным трубкам, сильно не изменяется, так как они состоят из живых клеток, которым необходимо поддерживать гомеостаз. Распределение передвигающихся веществ между корой и древесиной зависит также от обеспеченности растений водой. Во время засухи, когда устьица закрываются и транспирация резко тормозится, транспорт элементов минерального питания идет в основном по коре, когда воды достаточно, – по коре и древесине. Не только фосфор, но и большая часть других элементов минерального питания: калий, натрий, сера, хлор, магний, азот – также транспортируется по флоэме. Некоторые из них, например магний, для этого образуют комплексы с другими веществами. Поскольку железо при внутриклеточных значениях pH нерастворимо, оно, очевидно, должно перемещаться по растению в неионной форме или в форме хелатного железа. Многие микроэлементы, в том числе железо, марганец, цинк, молибден, в какой-то мере передвигаются по флоэме из зрелых тканей в незрелые. Элементы минерального питания передвигаются вверх по флоэме, когда происходит их реутилизация из листьев, о чем будет говориться позже. Итак, минеральные вещества, поглощенные из почвы, и продукты их первичной ассимиляции в корнях передвигаются как по сосудам, так и по ситовидным трубкам. Такое дублирование повышает надежность транспортной системы. Соотношение того и другого пути зависит от ряда условий: солевого состояния растения, обеспеченности его водой, концентрации питательного раствора и др. Не все ионы, попавшие в ксилему, достигают стебля или листьев. У крупных травянистых растений и деревьев состав ксилемного сока может значительно измениться во время его транспортировки из корней к листьям. Во-первых, в клеточных стенках ксилемы имеются многочисленные участки с отрицательными зарядами, способные связывать двухвалентные катионы и таким образом тормозить их транспорт по ксилеме. Часть ионов, например натрия, во время движения по ксилеме выходит из ксилемного сока и остается в клетках корня или стебля. По мере старения этот процесс активируется. При низкой температуре, наоборот, больше натрия поступает в побег. Во-вторых, ионы могут активно поглощаться из сосудов с помощью клеток камбия. Эти клетки регулируют количество и состав питательных веществ, передвигающихся по ксилеме. Если какого-то элемента слишком много в ксилемном соке, то он аккумулируется в клетках камбия; если мало, то выделяется из камбиальных клеток в ксилему. Элементы минерального питания могут транспортироваться не только вверх (восходящий ток), но и вниз по растению – нисходящий ток. Нисходящий транспорт зависит от температуры. Это является доказательством того, что он идет по ситовидным трубкам, так как восходящий ток, идущий по мертвым сосудам, нечувствителен к температуре. Во время транспорта минеральных веществ по стеблю может осуществляться обмен этими веществами между корой и древесиной. Механизм дальнего транспорта по сосудам – массовый ток, поэтому скорость движения элементов минерального питания по сосудам сильно зависит от интенсивности транспирации. По ситовидным трубкам ионы движутся со скоростью от 2–4 до 14 м/ч. Элементы, хотя и движутся вместе с транспираиионным током, поглощаются из сосудов более энергично не транспирирующими, а растущими частями растения. Направление движения веществ тоже определяется потребностями того или иного органа, интенсивностью его роста. Элементы двигаются в развивающиеся репродуктивные органы и плоды, оплодотворенные семяпочки, точки роста, т.е. туда, где идет энергичный рост и потребление ионов, синтез органических веществ. Благодаря энергичному потреблению ионов в этих органах и тканях в ксилемном соке сохраняется акропетальный градиент элементов минерального питания. Большое влияние на направление транспорта в растении оказывают гормоны: транспорт питательных веществ идет к тем органам, которые содержат больше гормонов, и в частности ауксинов. Как и поглощение, транспорт веществ зависит от интенсивности обмена веществ растения. Доказательством тому может служить суточная ритмичность движения пасоки. Даже в постоянных условиях днем пасока поднимается быстрее, чем ночью, причем корни работают ритмично и при удалении побегов. Ритмичность в деятельности корней сохраняется при разных условиях освещения, температуры и питания, но продолжительность периодов и особенно амплитуда колебаний изменяются при неблагоприятных условиях. Ритмичность в движении солей с пасокой говорит также о связи транспорта с работой живых клеток. Регуляция транспорта веществ происходит на всех уровнях организации живой материи. Механизмы ее различны: на клеточном уровне это открывание и закрывание ионных каналов, работа энергозависимых белков-переносчиков, на тканевом – отрывание и закрывание десмотубул и обмен информацией между клетками через апопласт, на организменном – изменение скорости обмена веществ.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
 Механизм транспорта ионов по симпласту до конца не выяснен. Предполагают, что это диффузия и массовый ток веществ из клетки в клетку по градиенту концентрации. Этот концентрационный градиент создают корневые волоски, активно поглощая вещества. Допускают возможность и поверхност-ной миграции ионов вдоль плазмодесм. В этом случае плазмалемма корневых клеток выполняет двойную функцию: поглощение (транспорт веществ через мембрану) и передвижение его вдоль своей внутренней поверхности, которая через плазмодесмы переходит из клетки в клетку. На движение ионов по симпласту влияет внутриклеточный транспорт, ускоряя диффузию вдоль ткани.
Механизм транспорта ионов по симпласту до конца не выяснен. Предполагают, что это диффузия и массовый ток веществ из клетки в клетку по градиенту концентрации. Этот концентрационный градиент создают корневые волоски, активно поглощая вещества. Допускают возможность и поверхност-ной миграции ионов вдоль плазмодесм. В этом случае плазмалемма корневых клеток выполняет двойную функцию: поглощение (транспорт веществ через мембрану) и передвижение его вдоль своей внутренней поверхности, которая через плазмодесмы переходит из клетки в клетку. На движение ионов по симпласту влияет внутриклеточный транспорт, ускоряя диффузию вдоль ткани.