
|
|
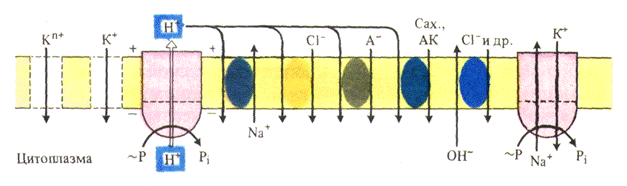
Поглощение элементов питания растениямиВ почвенном растворе ионы находятся в свободном состоянии или связаны с почвенными коллоидами. Поглощаются элементы минерального питания чаще всего в ионной форме: азот как NO3+ или NH4+, фосфор как НРО42- или Н2РО4-, сера как SO42-, молибден как МоО42-; калий, натрий, кальций, магний, тяжелые металлы (железо, марганец, медь) и цинк – в виде катионов; хлор – в виде хлорид-аниона; бор, вероятно, в форме недиссоциированной борной кислоты. Растение может поглощать и некоторые органические растворимые соединения, например аминокислоты. Однако основным источником питательных веществ являются минеральные соли. Длительное время считали, что сильно разбавленный почвенный раствор поступает в корни в неизмененном виде, поднимается по стеблю и потом «сгущается» в листьях в результате испарения воды, т.е. вещества поступают в растение в тех же количествах и соотношениях, в каких они находятся в почвенном растворе. Транспирационный ток считался одним из обязательных условий поступления веществ из почвы в корень, а затем в надземные органы. Однако более поздние опыты показали, что количество поступивших в растение и накапливаемых в нем минеральных веществ не пропорционально количеству прошедшей через него воды. В тропических лесах транспирация может быть подавлена из-за большой влажности воздуха. Несмотря на это, деревья достигают очень больших размеров и развивают большую листовую поверхность; следовательно, они обеспечивают себя всеми необходимыми элементами минерального питания. Изучение растений, выращенных в водной культуре, показало, что из очень разбавленных растворов соли поглощаются быстрее, чем вода, а из концентрированных растворов, наоборот, в растение быстрее поступает вода. Итак, минеральные соли и вода поступают в растение относительно независимо друг от друга и с помощью существенно различающихся механизмов. Однако независимость этих двух процессов не означает, что транспирационный ток не имеет никакого значения. Если в почвенном растворе много солей, то основная масса их передвигается из корней в надземные органы вместе с водой по сосудам ксилемы, т.е. вода облегчает транспорт веществ из корней в побеги (массовый ток). Если растение испытывает недостаток солей, то они передвигаются из корней в побеги не по древесине, а по коре, в этом случае транспирационный ток не может влиять на их транспорт. Не только различные соли, но даже анион и катион одной и той же соли поступают в растение из раствора с разной скоростью. Так, если в качестве источника азота используется сульфат аммония, то катион аммония поглощается растением более интенсивно, чем сульфат-анион, так как растению азот необходим в больших количествах, чем сера. В результате по мере роста растения в растворе, содержащем эту соль, может накапливаться серная кислота, повреждающая корни. Если в качестве источника азота используется нитрат натрия, то анион будет поступать в корни быстрее катиона. В окружающем растворе будет накапливаться NaHCО3. Эта соль, подвергаясь гидролизу, образует сильную щелочь NaOH и слабую кислоту Н2СО3, что вызывает подщелачивание питательного раствора. Азотнокислый аммоний является примером соли, у которой анион и катион поглощаются почти с одинаковой скоростью. Соль, у которой быстрее поглощается катион, называется физиологически кислой; у которой быстрее поглощается анион, – физиологически основной, у которой анион и катион поглощаются с одинаковой скоростью, – физиологически нейтральной. Растение поглощает вещества избирательно. Например, С4-растения поглощают большее количество калия, железа и кальция, чем C3-виды, произрастающие в тех же условиях. В результате избирательного поглощения соотношение поглощенных веществ в клетках может быть совсем другим, чем во внешнем растворе. Наблюдения показали также, что поглощение веществ идет не только избирательно, но и против градиента химического потенциала. Для такого поглощения нужно затратить энергию АТФ. Энергия дыхания может использоваться на перенос ионов против градиента химического потенциала и непосредственно, без предварительного запасания ее в АТФ. Согласно хемиосмотической теории П. Митчелла, в результате транспорта электронов по дыхательной цепи на наружной стороне внутренней митохондриальной мембраны накапливаются ионы водорода. При этом внутренняя сторона мембраны заряжается отрицательно. Катионы поступают внутрь органеллы, притягиваясь к отрицательно заряженной стороне мембраны. Таким образом, дыхательная цепь работает как протонный насос. Дыхательные яды, например динитрофенол, увеличивающие проницаемость мембраны для протонов, тормозят и поглощение ионов. Поглощение минеральных элементов в корневую систему растений может быть не только активным, но и пассивным. Пассивное поступление идет по градиенту химического потенциала. Однако главную роль в жизнедеятельности растения играет активное поглощение элементов. Как и поступление воды, процесс поглощения делят на два этапа: 1-й – поступление ионов из почвы или питательного раствора в свободное пространство клетки; 2-й – передвижение их из свободного пространства через плазмалемму в протопласт. Иногда поглощенные ионы могут транспорти-роваться из цитозоля в вакуоль или в другую органеллу. Механизмы поглощения веществ корнем. В свободное пространство клетки ионы поступают или из почвенного раствора, если речь идет об эпиблеме, или из свободного пространства соседних клеток. На первом этапе главными механизмами поглощения являются диффузия и адсорбция, на втором – мембранные транспортные белки, эндоцитоз. Клеточные стенки не могут служить преградой для поглощаемых веществ, потому что в них находятся межфибриллярные полости, через которые эти вещества свободно диффундируют. Ту часть объема корня, в которую минеральные вещества проникают или из которой выделяются путем свободной диффузии, называют апопластом. Анатомически апопласт представлен межфибриллярными полостями клеточных стенок и межклетниками. Объем свободного пространства составляет 5–10 % от всего объема корневой системы. Свободное пространство является внешним по отношению к протопластам клеток и внутренним по отношению к внешней среде. За счет диффузии вещества поступают из почвенного раствора в свободное пространство клетки (межфибриллярные полости), где концентрация их в основном меньше, чем в окружающем растворе. Скорость диффузии мала и уменьшается с увеличением ее продолжительности: за час вещество передвигается на 5 мм, за 24 ч – на 25 мм, а за год – на 500 мм. Поэтому диффузия не может играть важной роли в передвижении растворенных веществ на далекие расстояния, например из корня в лист. Итак, диффузия является основным механизмом поступления веществ в свободное пространство корня. Возможно, что через клеточную стенку вещества проходят не только с помощью диффузии, но и вместе с водой, когда, передвигаясь через мембрану, она захватывает одну или несколько небольших молекул растворенных в ней веществ (массовый ток). Стенки межфибриллярных полостей, составляющих свободное пространство клеток, имеют отрицательный электрический заряд, например, за счет карбоксильных групп пектиновых веществ, входящих в состав клеточных стенок. Поскольку вещества поглощаются главным образом в ионной форме, то на их поступление будет влиять и этот электрический заряд. Поэтому поступление ионов в свободное пространство клетки зависит не только от разности концентраций, но и от разностей электрических потенциалов. Существование отрицательного заряда на клеточной стенке облегчает поглощение катионов и затрудняет поглощение анионов.
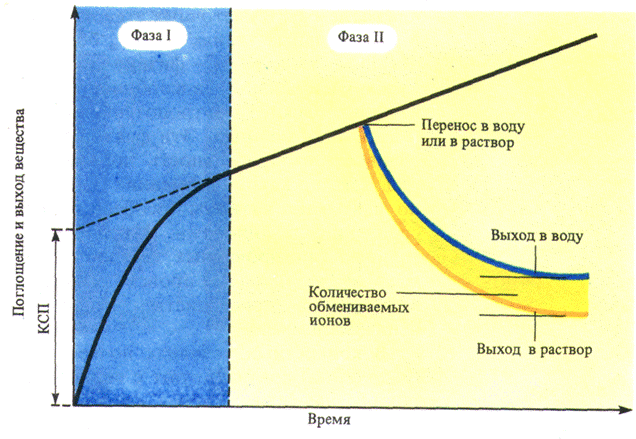
Кроме того, катионы могут адсорбироваться на клеточных стенках, поэтому концентрация катионов около стенок межфибриллярной полости больше, а анионов – меньше; в центре полости она равна концентрации катионов и анионов в наружном растворе. Из центра межфибриллярной полости ионы могут быть выделены в воду в результате диффузии. Ионы, адсорбированные на стенках межфибриллярных полостей, могут выделяться в солевой раствор путем обмена, причем адсорбированный ион вытесняется ионом, который содержится в солевом растворе в избытке (обменная адсорбция). Например, если корень, выдержанный ранее в растворе кальция, перенести в раствор калия, то кальций будет выделяться из корня в раствор. Если же корень поместить в дистиллированную воду, то этого не произойдет. Для явлений адсорбции характерна чрезвычайно большая начальная скорость, этим адсорбция отличается от диффузии. Вторым доказательством участия адсорбции в поглощении веществ служит существование состояния насыщения. Характерной особенностью поглощения является малая зависимость его вначале от температуры. Начальное поглощение зависит от величины pH. Особенно сильно концентрация водородных ионов влияет на соотношение количеств поглощаемых катионов и анионов. Итак, интенсивное поглощение вначале, существование состояния насыщения, независимость начального поглощения от температуры и полная зависимость от величины pH – все это доказывает, что в поглощении веществ принимает участие адсорбция. Благодаря высокой скорости, свойственной этому процессу, растение способно достаточно быстро адсорбировать на поверхности корня и затем накапливать в своих клетках необходимые ему питательные вещества из таких разбавленных растворов, какими обычно являются почвенные растворы. Войдя с помощью диффузии в свободное пространство корня, ионы адсорбируются не только на клеточных стенках, но и на плазмалемме, которая тоже несет электрический заряд. Разность электрических потенциалов на плазмалемме колеблется от 60 до 100, а иногда достигает 180 мВ, причем клетка заряжена отрицательно по отношению к наружному раствору. На втором этапе поглощения вещество из свободного пространства должно проникнуть в протопласт клетки. Для этого ему необходимо пройти через плазмалемму – основной барьер на пути диффузии ионов и молекул в клетку. Находящиеся в растворе в непрерывном хаотическом движении молекулы растворенного вещества при столкновении с этой мембраной либо отскочат от нее, либо адсорбируются на ней, либо пройдут через плазмалемму. В последнем случае говорят, что мембрана проницаема для данного вещества. Если вещество само пройти через плазмалемму не может, то помогают мембранные транспортные белки, а также эндоцитоз.
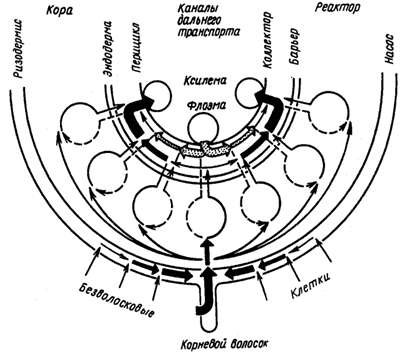
Разные механизмы работают не изолированно, а в различных сочетаниях, например «диффузия – адсорбция – переносчик» или «диффузия – адсорбция – ионный насос». В зависимости от природы поглощаемого вещества и условий в клетке удельное значение того или иного механизма изменяется. Организм сам регулирует, какой механизм поглощения должен работать. Хотя механизмы поглощения веществ делят на пассивные (диффузия, адсорбция) и активные (белки-переносчики, ионные каналы, эндоцитоз), это деление условно, так как для работы любого механизма нужна энергия. Например, для поддержания диффузии нужно, чтобы количество вещества в протопласте было меньше, чем в свободном пространстве клетки. Это возможно, если поглощаемое вещество сразу включается в обмен веществ, а для этого необходима энергия. Интенсивное поглощение клеткой веществ должно было бы привести к выравниванию концентраций и к прекращению поглощения. Однако этого не происходит, так как поступающие ионы, например нитрат-, сульфат- и фосфат-ионы, быстро включаются в клетке в органические соединения. Скорость включения ионов в обмен веществ в свою очередь определяет скорость их поглощения. Главный орган поглощения элементов минерального питания – корень. Удобрения можно вносить и через листья (внекорневая подкормка), но, как показали опыты с меченым фосфором, это вызывает их быстрое старение и опадение. В опытах с внекорневой подкормкой растения росли в 10 раз медленнее по сравнению с контрольными, получавшими такую же дозу фосфора через корни. Рассмотрим особенности строения корня как органа поглощения веществ. Эпиблема, покрывающая корень снаружи, непосредственно соприкасается с почвенным раствором и почвой. Поэтому первичное поступление ионов в корень происходитименно через клетки этой ткани. Эпиблема – основной барьер на пути поглощения ионов корнем. Это доказывается тем, что именно в клетках этой ткани как анионы, так и катионы накапливаются в наибольших количествах. Эпиблема как поглотительная ткань неоднородна: некоторые из ее клеток превращаются в корневые волоски. На 1 мм2 поверхности корня находится 200–400 корневых волосков; в результате поглощающая поверхность корня увеличивается в сотни раз. Корневой волосок – основной вход для ионов в корень; через другие клетки эпиблемы их поступает меньше. Косвенным доказательством этого служит и наибольшее число плазмодесм, выходящих из корневых волосков в клетки первичной коры. Первичная кора составляет до 86–90 % от всего объема корня, в ней много межклетников. Чем толще кора, тем больше общий объем корня и, следовательно, его поглощающая поверхность, так как в зоне корневых волосков корень имеет форму цилиндра.
Центральный цилиндр корня отделен от коры эндодермой, которая, регулирует переход веществ с апопластного пути на симпластный и обратно. Под эндодермой находится перицикл. Специально проведенный на электронно-микроскопических фотографиях подсчет показал, что в то время как между соседними клетками перицикла в клеточных стенках находится 45 тыс. плазмодесм, из каждой клетки перицикла в клетки центрального цилиндра идет только 12 тыс. Наблюдение позволило предположить, что в перицикле изменяется направление движения ионов с радиального на кольцевое. В результате из клеток перицикла ионы поступают непосредственно в сосуды, по крайней мере, у тех растений, у которых сосуды погружены в перицикл. Следовательно, роль перицикла можно сравнить с ролью кольцевой автодороги, позволяющей изменять направление движения. Функции находящихся в центре корня проводящих тканей хорошо известны: по ним поглощенные вещества транспортируются из корней в надземные органы. Корневая система неоднородна не только анатомически, но и физиологически. Разные зоны корня, отличающиеся разной скоростью роста, дыхания, поглощают вещества тоже с различной интенсивностью. Наиболее активно поглощают вещества клетки зоны растяжения и зоны корневых волосков. Опыты с проростками пшеницы показали, что корневые волоски не только увеличивают поверхность корня, но в их мембранах находятся транспортные белки, обладающие большей активностью по сравнению с такими же белками других клеток эпиблемы.
Если корень в течение какого-то времени поглощает вещества, то их концентрация около активно поглощающих участков корня уменьшится и скорость поглощения будет зависеть от скорости диффузии ионов в почве. У растущего кончика корня положение иное. Его клетки не только лучше поглощают, но благодаря делению и растяжению клеток кончик продвигается все дальше и дальше в новые участки почвы, где запасы необходимых веществ еще не истощились. В процессе роста корень двигается к питательным веществам. Поглощение в растущей части происходит намного быстрее, чем в других зонах корня, вблизи которых доставка веществ лимитирована. Чем быстрее ветвится корень, тем больше данная корневая система имеет зон растяжения и зон корневых волосков, т.е. зон активного поглощения. Итак, поглощение веществ зонами корня, закончившими рост, зависит лишь от скорости диффузии поглощаемых веществ; растущими зонами – от скорости диффузии и от скорости их роста. Мощное развитие корневых систем растений, особенно их мелких активных корешков и корневых волосков, беспрерывное распространение их во все новых и новых слоях почвы является необходимым условием поглощения веществ. Чем меньше подвижность данного иона в почве, тем большее значение приобретает мощность развития корневой системы, ее распространение в большем объеме почвы. Растение обладает большими потенциальными способностями к корнеобразованию. Изменяя схему внесения удобрений, можно управлять ростом корней в длину и скоростью их ветвления, следовательно, и урожаем. Кроме того, известно, что обрезка стимулирует ветвление корней, а это увеличивает количество зон с максимальным поглощением. Таким образом, быстрый рост корневой системы, стимулируя поглощение веществ, сам является одним из необходимых условий быстрого поглощения солей. Зоны корня отличаются между собой не только по скорости поглощения веществ, но и по интенсивности снабжения ими надземных органов. Так, клетки зоны деления и растяжения поглощают вещества исключительно для собственного («внутреннего») потребления. Они не только не транспортируют эти вещества в надземные органы, а, напротив, сами частично употребляют минеральные элементы, поглощенные в зоне корневых волосков. Это связано как с их функциями, так и с особенностями анатомии корня, в зоне растяжения которого проводящие ткани начинают закладываться. Таким образом, в корне различают зону, участвующую в поглощении питательных веществ (зона деления и растяжения клеток), и зону, участвующую как в поглощении, так и в снабжении питательными веществами надземных органов (зона корневых волосков). Однако неправильно было бы думать, что мощное развитие корней всегда является обязательным условием для удовлетворения потребностей растения в питательных веществах. Например, огромное количество сортов сахарного тростника с сильно развитой корневой системой низкоурожайны, поскольку много сахарозы расходуется на дыхание корней. Развитие растениями мощной и сильно разветвленной корневой системы является формой приспособления к поглощению малоподвижных веществ, к тому же часто весьма рассеянных в почве. При наличии в корнеобитаемой среде питательных веществ в легкодоступной форме и в достаточной концентрации мощное развитие корней не нужно, потому что корневая система работает обычно не с полной нагрузкой. Неравномерное распределение в почве питательных веществ привело к тому, что у корней в процессе эволюции выработалась способность быстрее расти в том направлении, где больше концентрация недостающего элемента. Это свойство получило название хемотропизма. Итак, скорость и направление движения корня в почве, величина поверхности, активно поглощающей соли и снабжающей ими другие органы, скорость включения ионов в метаболизм обусловлены ростом корня. Следствием этой зависимости является то, что при недостатке питательных веществ прежде всего тормозится рост побегов, а рост корней в длину, наоборот, стимулируется, что позволяет корню быстрее пройти слой почвы, бедный питательными веществами. Значение изучения закономерностей формирования корневых систем и поглощения ими элементов минерального питания важно, во-первых, для решения некоторых вопросов агротехники (глубины пахотного слоя, глубины заделки семян и удобрений, выбора методов обработки почвы и орошения); во-вторых, существуют определенные корреляции между развитием корней и приспособляемостью растений к засухе, к повышенной влажности или к повреждению вредителями.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
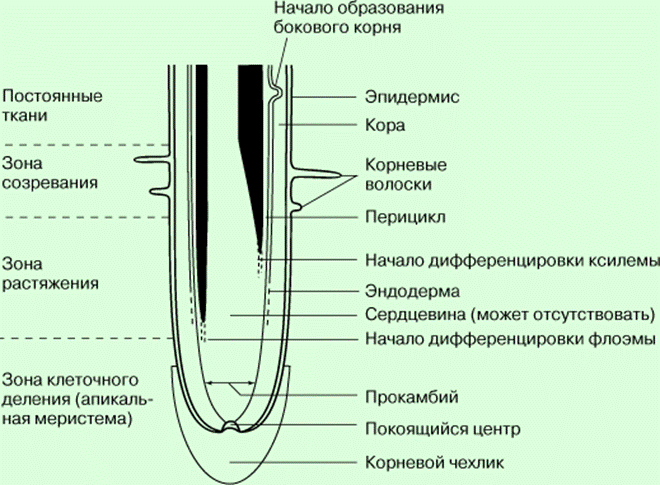
 В этих зонах наиболее активно происходит синтез белков и других компонентов протопласта, являющихся акцепторами ионов, поглощаемых из внешней среды. В зоне опробковения поглощение ионов на единицу поверхности корня резко уменьшается. Зона наиболь-шего поглощения, зависящая от особенностей растения, почвы, уровня грунтовых вод, орошения и распреде-ления элементов минераль-ного питания, с возрастом растения опускается вниз. Итак, в каждом корне существует значительный градиент поглотительной способности, который уменьшается от кончика корня к его базальной части.
В этих зонах наиболее активно происходит синтез белков и других компонентов протопласта, являющихся акцепторами ионов, поглощаемых из внешней среды. В зоне опробковения поглощение ионов на единицу поверхности корня резко уменьшается. Зона наиболь-шего поглощения, зависящая от особенностей растения, почвы, уровня грунтовых вод, орошения и распреде-ления элементов минераль-ного питания, с возрастом растения опускается вниз. Итак, в каждом корне существует значительный градиент поглотительной способности, который уменьшается от кончика корня к его базальной части.