
|
|
ЛИНИИ МЫШЕЙ С ГЕНЕТИЧЕСКИМИ ДЕФЕКТАМИ, ЗАТРАГИВАЮЩИМИ ИММУННУЮИНБРЕДНЫЕ ЖИВОТНЫЕ Для проведения фундаментальных исследований в иммунологии лучший объект - инбредные мыши. Инбредные животные - это животные, полученные путем инбридинга (in breed - выводить породу, разводить), т.е. последовательных близкородственных скрещиваний с целью получения гомозиготного и генетически идентичного потомства. Среди потомков для дальнейших скрещиваний сначала отбирают особей по признакам внешнего сходства, в последующих поколениях уже тестируют на совпадение групп крови и приживление кожных лоскутов. Через 20 поколений и более такой селекции получают мышей с весьма высокой степенью гомозиготности, обозначаемых как чистая линия, в пределах которой все животные генетически почти идентичны (например, как однояйцевые близнецы у человека). Главная цель выведения чистых линий мышей и исследований на них - получение возможности многократного повторения экспериментов на генетически одинаковых организмах, т.е. обеспечение воспроизводимости результатов исследований в высоком смысле этого понятия, что полностью исключено при решении многих иммунологических задач с использованием беспородных животных. Подобные проблемы существуют при оценке результатов иммунных процессов у человека. Мыши стали исключительными экспериментальным животными в иммунологии в силу ряда причин, главные из которых следующие: 1) короткий срок беременности (21 сут) и множественное потомство от каждой самки (5-8 детенышей в одни роды) позволяют весьма быстро вывести чистые линии, что важно по вышеназванным причинам; 2) себестоимость содержания мышей по сравнению с таковой других млекопитающих наименьшая; 3) структура и функция иммунной системы мыши и человека во многом сходны; 4) выведение чистых линий мышей показало, что, например, некоторые из них (несмотря на гомозиготность) весьма крепкие и здоровые, т.е. не всякий инбридинг приводит к вырождению. Кроме того, путем целенаправленного отбора тех или иных свойств созданы многочисленные линии мышей с точно заданными характеристиками, и это позволяет выбирать особей, необходимых для достижения конкретных научных целей. Характеристики животных разных линий занесены в соответствующие документы; на них ориентируются питомники по разведению чистолинейных мышей, имеющиеся во всех странах, где успешно занимаются проблемами экспериментальной иммунологии. Из наиболее прославленных питомников хотим упомянуть Джексоновскую лабораторию (The Jackson Laboratory) в США. Ежегодно она поставляет в университеты, медицинские институты и научно-исследовательские лаборатории всего мира приблизительно 2 млн животных 2500 разных линий, стоков и животных-моделей. Около 97% этих животных можно приобрести только в Джексоновской лаборатории. В каждом питомнике разводимые и поддерживаемые линии мышей имеют паспорт, систематизированы в соответствующих базах данных и доступны для широкого применения. Известен гаплотип (Н-2) мышей разных линий, их окрас, поведенческие характеристики, особенности функционирования иммунной системы и прочие свойства, необходимые не только для иммунологических исследований, но и исследований в других областях биологии и медицины (онкология, фармакология, экология и т.д.). Мы приводим характеристику некоторых наиболее известных линий мышей, которые экспериментаторы выбирают с теми или иными определенными целями (табл. 1.1). Таблица 1.1.Линии инбредных мышей, наиболее часто применяемые в исследованиях (Кондратьева И.А., Ярилин А.А., 2004)
C57B1/6(рис. 1.2, см. также цв. вклейку) -черного цвета мыши, подвижные, агрессивного поведения. Balb/c(рис. 1.3) -белые мыши с хрупким здоровьем. Однако это самая востребованная линия для гибридомной биотехнологии, потому что линии миелом, на основе которых получают гибридомы, ведут свою «родословную» от перевивной линии лейкозных клеток МОРС- 21, происходящей от мышей Balb/c. Гибридомы хорошо растут в брюшной полости живых сингенных мышей в виде асцитных опухолей. Кроме собственно чистых линий мышей, генетики научились выводить так называемых конгенныхмышей. Так называют линии, отличающиеся друг от друга небольшой областью генома (иногда одним геном).
В основе выведения конгенных линий мышей лежит генетический прием возвратного скрещивания - получение потомства в ряду поколений от скрещивания гетерозиготы (потомков гомозиготных родителей, генетически отличающихся между собой) с одним из исходных гомозиготных родителей. Смысл подобного скрещивания - внедрить комплекс Н-2 донорской линии А в генотип основной линии В. На рисунке 1.4 представлены донорская маркирующая линия А и основная линия В. От скрещивания гомозиготных особей этих двух линий получают гибриды первого поколения F1, (a/b; генерация 1). При дальнейшем скрещивании гибридов F1 с особями основной линии В получают потомство, состоящее как из гомозигот (b/b), так и гетерозигот (а/b) по комплексу Н-2. В последующих скрещиваниях отбираются только гетерозиготные особи, имеющие признак «а» (Н-2а), который определяется по приживлению кожного трансплантата от маркирующей линии A и положительной серологической реакции клеток крови с анти-А-сывороткой. По мере продолжения скрещиваний а-положительных особей с особями основной линии В доля генома линии А постоянно снижается, но при этом для дальнейшего размножения из потомства отбирают только тех особей, которые сохраняют признак «а» (H-2a). К двенадцатому поколению (генерация ? 12) практически весь геном отбираемых после гибридизации мышей представлен основной линией В, за исключением признака «а», по которому шел отбор. Дальнейшая задача состоит в переводе признака «а» в гомозиготное состояние. Для этой цели гетерозигот (а/b) скрещивают между собой и отбирают для дальнейшего размножения только тех особей из потомства, которые отторгают кожный трансплантат, взятый от особей линии В, и не дают реакции с анти-В-сывороткой. Подобный отбор выявляет особей с отсутствием признака «b» (H-2b) и гомозиготность по признаку «а» (H-2a). Таким образом, в результате применения данной схемы скрещивания в геном основной линии В внедряется комплекс Н-2 маркирующей линии А (рис. 1.4). С момента перевода комплекса H-2a в гомозиготное состояние констатируется получение новой конгенной (по отношению к основной) линии В (Klein J., 1975). ЛИНИИ МЫШЕЙ С ГЕНЕТИЧЕСКИМИ ДЕФЕКТАМИ, ЗАТРАГИВАЮЩИМИ ИММУННУЮ СИСТЕМУ SCID(англ. severe combined immunodeficiency) - мыши, страдающие тяжелым иммунодефицитом в результате мутации в генах RAG, ответственных за перегруппировку генов иммуноглобулинов и Т-клеточного рецептора. Животные практически лишены Т- и В-лимфоцитов и могут жить в безмикробных условиях, но необязательно - в полностью стерильных. Эти мыши не отторгают ксеногенные ткани, в частности им можно вводить с расчетом на приживление самые разнообразные клетки человека. В 1959 г. Расселом и соавт. описаны определенные частично инбредные мыши - спонтанные мутанты-самцы, которые вскоре после рождения покрывались чешуйчатой перхотью (рис. 1.5), заметно отставали в росте, страдали от тяжелой диареи и умирали в возрасте около 3 нед. При морфологическом исследовании у мышей наблюдали массивную лимфоаденопатию, спленомегалию, аномаль-
ную инфильтрацию лимфоцитами кожи, печени, легких, эндокринных желез. Мутантных самцов назвали «скуpфи» (англ. scurfy - покрытые перхотью). Только в 2001 г. M.E. Brunkow и соавт. показали: мутация scurfy затронула ген фактора транскрипции FoxP3 (вставка двух пар оснований в 8-й экзон этого гена), локализованного на Х-хромосоме. В том же 2001 г. несколько исследователей (C. Bennett, R. Wildin) показали: у детей с синдромом IPEX поврежден тот же ген (Xp11.23-Xq13.3), что и у мышей scurfy - FoxP3. В 2003-2005 гг. Джейсон Фонтенот и Александр Руденский показали: экспрессия гена FoxP3 является существенным и определяющим моментом в дифференцировке в тимусе Т-регуляторных клеток (Treg), открытых Сакагучи в 1995-1996 гг. и ответственных за поддержание иммунологической толерантности к нормальным тканям своего организма и гомеостаза в иммунной системе, а также за контроль аутоиммунных и опухолевых заболеваний.
Nude(лишенные волосяного покрова) - замеченные и отобранные в 1960 г. мыши со спонтанной мутацией, в результате которой у мышей-гомозигот по данной мутации (nu/nu) отсутствуют тимус и волосяной покров (рис. 1.6, см. также цв. вклейку). Мутантный ген поддерживают при размножении мышей в гетерозиготном состоянии; он перенесен мышам нескольких других линий, например Balb/c, CBA/Ca, C57Bl/10ScSn и пр.
C57Bl/6-bg/bg (англ. beige - бежевый) - мыши-мутанты бежевого окраса из исходно черной линии. Характеризуются существенно сниженной активностью NK-клеток и фагоцитов с повреждением лизосомальных структур. AKR - белые мыши (рис. 1.7, см. также цв. вклейку). У 90% особей обоего пола к возрасту 6-8 мес развиваются «спонтанные» тимомы и лейкоз.
W/Wv - мыши с существенным дефицитом тучных клеток в слизистых оболочках. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
 Продолжение табл. 1.1
Продолжение табл. 1.1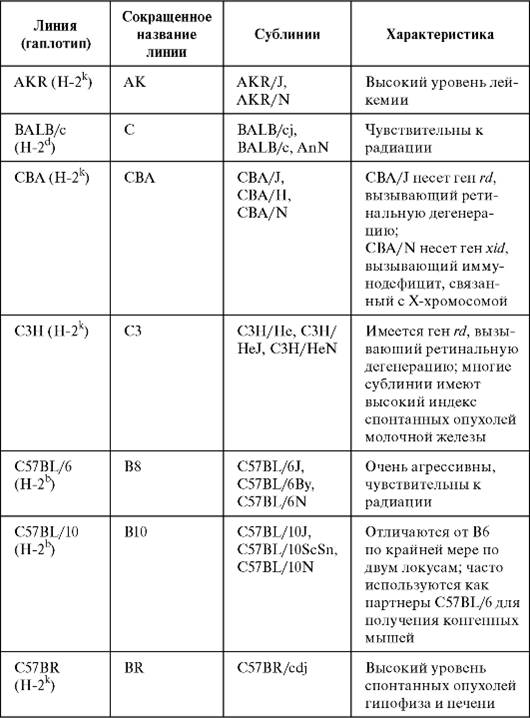 Продолжение табл. 1.1
Продолжение табл. 1.1 Окончание табл. 1.1
Окончание табл. 1.1 СВА/J(рис. 1.1, см. также цв. вклейку) и гибриды первого поколения (CBA/J×C57Bl.6)F1 -серо-бурые здоровые выносливые мыши, которые хорошо переносят облучение в кроветворных летальных дозах, и по этой причине их часто используют в радиационных моделях.
СВА/J(рис. 1.1, см. также цв. вклейку) и гибриды первого поколения (CBA/J×C57Bl.6)F1 -серо-бурые здоровые выносливые мыши, которые хорошо переносят облучение в кроветворных летальных дозах, и по этой причине их часто используют в радиационных моделях. Рис. 1.1.Мышь линии СВА/J
Рис. 1.1.Мышь линии СВА/J Рис. 1.2.Мышь линии C57B1/6
Рис. 1.2.Мышь линии C57B1/6 Рис. 1.3.Мышь линии Balb/c
Рис. 1.3.Мышь линии Balb/c Рис. 1.4.Схема получения конгенных линий (Галактионов В.Г., 1986)
Рис. 1.4.Схема получения конгенных линий (Галактионов В.Г., 1986) Рис. 1.5.Мышь линии Scurfy («покрытые перхотью»)
Рис. 1.5.Мышь линии Scurfy («покрытые перхотью») Рис. 1.6.Мышь линии Nude
Рис. 1.6.Мышь линии Nude Рис. 1.7.Мышь линии AKR/J
Рис. 1.7.Мышь линии AKR/J