|
|
Материнские мРНК и дроблениеДробление у морского ежа требует непрерывного синтеза белка на запасенных материнских мРНК. Если яйца морского ежа оплодотворяются в присутствии ингибитора трансляции, то дробления не происходит, хотя оба пронуклеуса сливаются. Кроме того, не образуется митотического веретена, хромосомы не конденсируются и ядерная мембрана не разрушается (Wagenaar, Mazia, 1978).
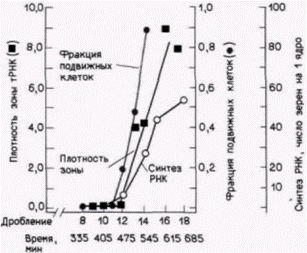
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. 216_______________ ГЛАВА 14______________________________________________________________________________ Таким образом, некоторые белки, кодируемые материнскими мРНК, играют, по-видимому, важную роль в делении клеток при дроблении. Эта гипотеза была подтверждена открытием группы белков, названных циклинами (Evans et al., 1983). Циклины кодируются материнскими мРНК и не обнаруживаются в неоплодотворенной яйцеклетке. После оплодотворения их синтез резко возрастает. Особенно замечательно в циклинах то, что они разрушаются при клеточном делении и после завершения каждого деления дробления ресинтезируются заново на запасенных мРНК (рис. 14.22). Синтез циклинов снижается, когда зародыш завершает стадию бластулы. Впервые циклины были обнаружены у морского ежа, позднее у других видов. Белки А и В у двустворчатого моллюска Spisula (о чем упоминалось ранее) представляют собой циклин, и их активность коррелирует с клеточным циклом. После оплодотворения эти белки непрерывно синтезируются на запасенных материнских мРНК и накапливаются в течение S-фазы и ранней М-фазы клеточного цикла. В ходе митоза эти белки разрушаются. Полагают, что циклины могут определять время клеточных делений, так как инъекция циклина А в ооциты Xenopus стимулирует их к вхождению в мейоз (Swenson et al., 1986). Дополнительные сведения и гипотезы: Активация генома зародыша Скорости развития животных различных видов чрезвычайно сильно варьируют. К 24 часам после оплодотворения личинка дрозофилы уже вылупилась из яйца и интенсивно питается, зародыши амфибий находятся на стадии поздней гаструлы или ранней нейрулы, а зародыши морского ежа – на стадии поздней бластулы или ранней гаструлы, содержащей сотни клеток. Зародыши млекопитающих не торопятся. К 24 часам после оплодотворения зигота мыши разделилась всего лишь один раз, а яйцеклетку человека отделяют от первого деления дробления примерно 6 ч. Исходя из такого разнообразия, можно ожидать, что геномы зародышей различных видов активируются в разное время (табл. 14.4). У зародышей Xenopus резкая активация наблюдается после двенадцатого деления дробления (Newport, Kirschner, 1982). До этого времени ядерная транскрипция у нормальных зародышей не обнаруживается, но после перехода к средней бластуле транскрипция начинается с высокой скоростью (рис. 14.23). У морского ежа, по-видимому, ядра зародыша функционируют постоянно. Даже в пронуклеусах перед их слиянием наблюдается транскрипция (в том числе синтез новых гистоновых мРНК), хотя уровень ее очень невысок (Poccia et al., 1985). Эти новосинтезированные мРНК присоединяются к большому фонду материнских мРНК. В ходе первых четырех дроблений хроматин образуется преимущественно из гистонов, запасенных в цитоплазме ооцита, и из гистонов, синтезированных на материнских мРНК. Однако, начиная со стадии 16 клеток большая часть гистонов синтези-
Таблица 14.4. Активация геномов зародышей и продолжительность функционирования материнской мРНК
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с. __________________ ТРАНСЛЯЦИОННАЯ И ПОСТТРАНСЛЯЦИОННАЯ РЕГУЛЯЦИЯ РАЗВИТИЯ____________________ 217
руется на молекулах мРНК, которые были транскрибированы в ядрах клеток зародышей (Goustin, 1981). Эта картина существенно отличается от той. что наблюдается у зародышей Xenopus, у которых значительный фонд запасенных в ооците гистонов и большой запас ооцитных гистоновых мРНК используются тысячами клеток. У дрозофилы транскрипция в ядре наблюдается впервые после его десятого деления. Это первый клеточный цикл с G2-фазой, и длительность G2-фазы увеличивается с 10 мин после 10-го цикла до 60 мин после 14-го цикла. В G2-фазе 14-го цикла уровень транскрипции генома наибольший в течение всего эмбриогенеза (Anderson, Lengyel, 1979; Weir, Kornberg, 1985). Ядра дрозофилы приобретают компетентность к транскрипции на 10-й цикл, но большинству генов для активации требуется более продолжительная G2-фаза (Edgar, Schubiger, 1986). Высокая транскрипционная активность, свойственная зародышам 14-го цикла, может быть индуцирована ранее с помощью искусственного удлинения G2-периода циклогексимидом. Эту активацию можно осуществить в зародышах 10-го цикла, но не в более ранних. Следовательно, большинство генов приобретают способность к активации в течение 10-го цикла, но не инициируют свою транскрипцию вплоть до 14-го цикла.
В отличие от организмов, обсуждавшихся выше, для мышей характерен определенный период, когда используются материнские мРНК, за которым следует период, когда эти мРНК заменяются на ядерные транскрипты. Материнские мРНК существуют около двух суток - примерно такое же время они существуют у представителей других эволюционных групп, и затем, в течение вторых суток, материнские мРНК быстро разрушаются (Clegg, Piko, 1983). Когда продукты, кодируемые материнскими мРНК. распадаются, они заменяются новыми белками, синтезированными на мРНК, новообразованной в ядре. В большинстве случаев хромосомы, полученные от спермия, активируются, вероятно, одновременно с хромосомами, полученными от яйца (Gilbert, Solter, 1985). У всех исследованных видов животных существует период, когда события раннего развития контролируются посредством мРНК и белков, запасенных в цитоплазме ооцита. У большинства видов (млекопитающие составляют исключение) ядерный геном активируется задолго до того, как разрушаются материнские мРНК, поэтому оба набора мРНК транслируются одновременно. В конечном счете, когда на первые или вторые сутки материнские мРНК разрушаются, транскрипты с генома зародыша начинают играть главную роль. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|