|
|
Детерминация положения бластопораОписав в основных чертах гаструляцию у амфибий, мы можем теперь рассмотреть ее более подробно. Гаструляция в жизни животного не существует как независимый процесс. Подготовка к ней может быть прослежена буквально до момента слияния спермия с яйцом. Неоплодотворенное яйцо обладает анимально-вегетативной полярностью (рис. 2.36, 2.38). На анимальном полюсе пигментированная кортикальная цитоплазма располагается над светлой областью цитоплазмы, содержащей материал, который ранее находился в ядре ооцита. (На стадии метафазы первого деления мейоза этот материал высвобождается вблизи того места, где образуются полярные тельца.) Постепенно этот материал сливается с областью, содержащей большие количества рибосом и митохондрий: ниже находится область цитоплазмы с гранулами гликогена и мелкими желточными пластинками. Вегетативная часть неоплодотворенного яйца содержит более крупные желточные пластинки и окружена светлой кортикальной цитоплазмой.
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. ГАСТРУЛЯЦИЯ: ПЕРЕРАСПРЕДЕЛЕНИЕ КЛЕТОК 121
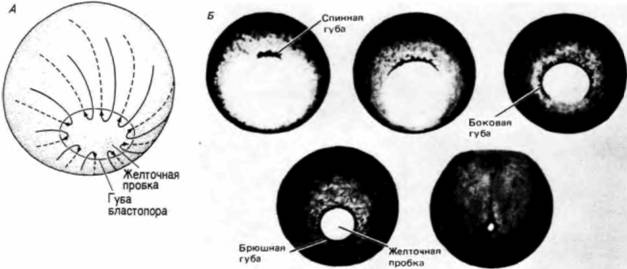
Судьба этих областей может быть предсказана еще до оплодотворения. Поверхность анимального полушария станет клетками эктодермы (эктодермальный эпителий и нервная система), поверхность вегетативного полушария даст клетки энтодермы (кишечный эпителий и связанные с ним органы), а мезодермальные клетки будут формироваться из внутренней цитоплазмы, лежащей вокруг экватора. Следовательно, зародышевые листки могут быть в общих чертах картированы в неоплодотворенном яйце: но это еще ничего не говорит нам о том, из какой части яйца образуется брюшная часть, а из какой - спинная. Переднезадняя (голова—хвост) и дорсовентральная (спина — живот) оси еще не детерминированы. Дорсовентральную и переднезаднюю оси детерминирует смещение цитоплазмы зиготы при оплодотворении. В гл. 2 мы обсуждали поворот кортикального слоя цитоплазмы относительно внутренней цитоплазмы яйца у лягушки. Внутренняя цитоплазма остается в прежнем положении из-за большого количества желтка в ней, тогда как кортикальный слой цитоплазмы активно смещается на 30о по отношению к своему первоначальному положению. Этот поворот приводит к тому, что анимально-вегетативная полярность наружного слоя яйца смещается на 30о по отношению к анимально-вегетативной полярности внутренней цитоплазмы. Таким образом возникает новое состояние симметрии. Вместо радиальной симметрии устанавливается двусторонняя, или билатеральная (рис. 4.17). Та область внутренней цитоплазмы вегетативного полушария, которая теперь лежит под кортикальным слоем цитоплазмы анимального полушария, по-видимому, активизируется и приобретает способность инициировать гаструляцию. Обычно это происходит на стороне, где образуется «серый серп», отстоящий на 180о от места проникновения спермия (рис. 4.17). Сторона, на которой спермий проникает в яйцо, станет брюшной поверхностью зародыша: противоположная сторона, где начинается гаструляция, будет спинной поверхностью зародыша (Gerhart et al.. 1981: Vincent et al., 1986; Gerhart et al., 1986). Для индукции этих движений цитоплазмы яйца спермий не является необходимым. Если искусственно стимулировать дробление энуклеированного яйца, то смещения кортикального слоя в положенное время все же происходит, однако направление его непредсказуемо. (В яйцах, оплодотворенных двумя спермиями, кортекс поворачивается только в одном направлении.) Спермий, по-видимому, дает сигнал для ориентации автономного смещения цитоплазмы в пространстве. Однако смещение цитоплазмы очень существенно: если оно блокировано, то дорсальные осевые структуры не формируются. Направление движения цитоплазмы определяет, какая сторона яйца должна стать спинной, а какая — брюшной (Gerhart et al., 1986; Vincent. Gcrhart, 1987). Направление смещения, определяемое местом проникновения спермия, может быть подавлено путем механического изменения пространственных со-
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 122 ГЛАВА 4
отношений между кортикальной и субкортикальной цитоплазмой. Это может быть достигнуто таким поворотом яйца, в результате которого внутренние компоненты попадают в разные области яйца (Pfluger. 1883; Penners. Schleip. 1928: Pasteels. 1948; Gerhart et al.. 1981). В норме оплодотворенное яйцо ориентировано таким образом, что тяжелые желточные пластинки в вегетативном полушарии в соответствии с действием силы тяжести располагаются в нижней части яйца. Если воспрепятствовать повороту яйца, погрузив его в раствор полисахарида (чтобы обезво-
дить перивителлиновое пространство между яйцом и оболочкой оплодотворения), то затем яйцо можно повернуть набок на 90о, так чтобы анимально-вегетативная ось приняла вместо вертикального горизонтальное положение, а место проникновения спермия было обращено кверху (Gerhart et al.. 1981; Kirschncr, Gcrhart. 1981; Cooke. 1986). Если оплодотворенное яйцо остается в таком положении в течение 30 мин. начиная с середины первого цикла деления дробления, то почти у всех зародышей гаструляция начинается на той же стороне, на которой спермий проник в яйцо (рис. 4.18). Роль нормального смещения цитоплазмы можно продемонстрировать в опытах с подавлением этого смещения и последующим наблюдением за зародышами, развивавшимися из таких яиц. Воздействия, вызывающие деполимеризацию микротрубочек (холодовой шок, высокое гидростатическое давление или погружение яиц в нокодазол), блокируют поворот цитоплазмы (Mânes et al.. 1978). Обработанные таким образом яйца дробятся, но никогда не гаструлируют нормально и не приобретают дорсовентральной полярности. Гаструляция у них протекает симметрично, и они превращаются в трехслойный цилиндр, лишенный спинных компонентов, таких, как сомиты и центральная нервная система. Эти зародыши погибают после того, как будут исчерпаны их запасы желтка (Gerhart et al., 1986). Однако если эти яйца были повернуты на 90о за 40 мин до первого деления дробления (тем самым имитировался нормальный поворот цитоплазмы), то у зародышей формировались нормальные осевые структуры, а гаструляция начиналась на той стороне, которая должна была стать спинной (Sharf, Gerhart, 1983). Перераспределение цитоплазмы в течение первого цикла делений дробления является необходимым условием для нормальной гаструляции. Как и следовало ожидать из предыдущего
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. ГАСТРУЛЯЦИЯ ПЕРЕРАСПРЕДЕЛЕНИЕ КЛЕТОК 123
обсуждения, влияние силы тяжести оказалось возможным дублировать центрифугированием яиц в течение короткого времени (Black, Gerhart. 1985). Оплодотворенные яйца помешали в желатин для их иммобилизации и поворачивали так, чтобы место проникновения спермия в яйцо было либо удалено от ротора центрифуги, либо обращено к нему. Затем эти яйца подвергали слабому вращению (10 — 50 g) в течение 4 мин. Если воздействие оказывали в первые две трети (70%) периода первого цикла делений дробления, то смещение цитоплазмы, вызванное центрифугированием, преобладало над смещением цитоплазматических компонентов, индуцированным проникновением спермия. По прошествии этого времени (т.е. 70%) цитоплазма становится более ригидной и ее смещения под влиянием центрифугирования не наблюдается. Кроме того, если яйца подвергали четырехминутному центрифугированию при 30 g в период, равный 40—50% времени прохождения первого клеточного цикла, то возникало два места начала гаструляции, что приводило к развитию частично сросшихся близнецов (рис. 4.19). Блэк и Герхарт (Black, Gerhart, 1986) высказали гипотезу, что образование близнецов вызвано формированием двух областей взаимодействия: одна ось формируется там, где смещенная центрифугированием цитоплазма взаимодействует с лежащими над ней компонентами: другая формируется после того, как цитоплазма, смещающаяся под влиянием силы тяжести, возвращается в свое первоначальное положение, где она реагирует повторно. Близнецы могут также образоваться в нормальных гравитационных условиях, если яйцо поместить местом проникновения спермия кверху, освободив его при этом от оболочки оплодотворения (Gerhart et al., 1981). Возможность влиянием силы тяжести и центрифугированием вызвать образование двух осевых структур в яйцах, вполне нормальных в других отношениях, и, напротив, вызвать образование осевых структур в яйцах, у которых подавлено нормальное смещение цитоплазмы, убедительно свидетельствует о том, что вызванные этими воздействиями внутренние смещения замещают процесс, в норме участвующий в определении места, на котором начнется гаструляция. Способность зародыша к образованию двух функциональных губ бластопора позволяет утверждать, что область серого серпа (где обычно начинается гаструляция) не является чем-то уникальным. Факторы, индуцирующие гаструляцию, по-видимому, возникают в результате взаимодействия между анимальной и вегетативной цитоплазмой; это взаимодействие, вероятно, активирует некий компонент в субкортикальной вегетативной цитоплазме. Гимлих и Герхарт (Gimlich. Gerhart, 1984) провели серию опытов с трансплантацией, подтвердивших гипотезу о том, что фактор или факторы, инициирующие гаструляцию, первоначально содержатся в глубокой цитоплазме, а не в кортексе серого серпа. Эти авторы показали, что у зародыша шпорцевой лягушки на стадии 64 бластомеров три наиболее дорсально расположенных вегетативных бластоме-
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. 124 ГЛАВА 4
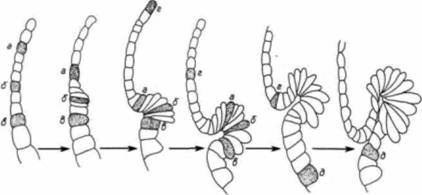
pa способны индуцировать образование спинной губы бластопора и полного комплекса осевых структур у облученных ультрафиолетовым светом реципиентов (которые иначе не могли бы гаструлировать) (рис. 4.20, А). Кроме того, эти три бластомера, лежащие под областью проспективной спинной губы бластопора. могут также индуцировать вторичную инвагинацию и образование осевых структур, будучи трансплантированы на брюшную сторону нормального необлученного 64-клеточного зародыша (рис. 4.20, Б). Следовательно, эта маленькая группа вегетативных бластомеров ответственна за появление в прилегающих к ним краевых клетках способности инвагинировать и формировать дорсальные мезодермальные структуры зародыша. Представляется вероятным, что внутреннее перемещение цитоплазмы, ориентация которого в норме определяется проникновением спермия, является причиной асимметричного распределения субклеточных факторов. Эта асимметрия создает в яйце дорсовентральные различия, которые в конечном счете определяют положение бластопора. Кроме того, происходит прогрессивное накопление какого-то фактора (или факторов), необходимого для формирования спинной губы бластопора. Малацински и др. (Malacinski et al., 1980) не обнаружили в дорсальной области оплодотворенного яйца и на ранних стадиях дробления у амфибий активности, необходимой для формирования спинной губы бластопора. Однако такая активность в этой области выявлялась на стадиях средней и поздней бластулы. Ее можно было обнаружить и в вегетативных бластомерах, лежащих в наиболее дорсальном участке 64-клеточного зародыша лягушки. Следовательно, факторы, ответственные за формирование бластопора, по-видимому, локализуются в области, где будет за-
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с. _______________ ГАСТРУЛЯЦИЯ: ПЕРЕРАСПРЕДЕЛЕНИЕ КЛЕТОК 125
кладываться бластопор, лишь незадолго до начала гаструляции. Эти вегетативные клетки затем взаимодействуют с клетками, лежащими непосредственно над ними (в области серого серпа), побуждая их гаструлировать и формировать дорсальные осевые структуры. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|