|
|
Строение, развитие и функциональное значение различных отделов центральной нервной системыСпинной мозг Спинной мозг представляет собой длинный тяж. Он заполняет полость позвоночного канала и имеет сегментарное строение, соответствующее строению позвоночника. В центре спинного мозга расположено серое вещество — скопление нервных клеток, окруженное белым веществом, образованным нервными волокнами (см. рис. 68). В спинном мозге находятся рефлекторные центры мышц туловища, конечностей и шеи. С их участием осуществляются сухожильные рефлексы в виде резкого сокращения мышц (коленный, ахиллов рефлексы), рефлексы растяжения, сгибательные рефлексы, рефлексы, направленные на поддержание определенной позы. Рефлексы мочеиспускания и дефекации, рефлекторного набухания полового члена и извержения семени у мужчин (эрекция и эякуляция) также связаны с функцией спинного мозга. Спинной мозг осуществляет и проводниковую функцию. Нервные волокна, составляющие основную массу белого вещества, образуют проводящие пути спинного мозга. По этим путям устанавливается связь между различными частями ЦНС и проходят импульсы в восходящем и нисходящем направлениях. По этим путям поступает информация в вышележащие отделы мозга, от которых отходят импульсы, изменяющие деятельность скелетной мускулатуры и внутренних органов. Деятельность спинного мозга у человека в значительной степени подчинена координирующим влияниям вышележащих отделов ЦНС. Обеспечивая осуществление жизненно важных функций, спинной мозг развивается раньше, чем другие отделы нервной системы. Когда у эмбриона головной мозг находится на стадии мозговых пузырей, спинной мозг достигает уже значительных размеров. На ранних стадиях развития плода спинной мозг заполняет всю полость позвоночного канала. Затем позвоночный столб обгоняет в росте спинной мозг, и к моменту рождения он заканчивается на уровне третьего поясничного позвонка. У новорожденных длина спинного мозга 14—16 см, к 10 годам она удваивается. В толщину спинной мозг растет медленно. На поперечном срезе спинного мозга детей раннего возраста отмечается преобладание передних рогов над задними. Увеличение размеров нервных клеток спинного мозга наблюдается у детей в школьные годы.
Головной мозг
Головной мозг состоит из трех основных отделов — заднего, среднего и переднего мозга, объединенных двусторонними связями. Задний мозг является непосредственным продолжением спинного мозга. Он включает продолговатый мозг, мост и мозжечок. Продолговатый мозг играет значительную роль в осуществлении жизненно важных функций. В нем расположены скопления нервных клеток — центры регуляции дыхания, сердечно-сосудистой системы и деятельности внутренних органов. На уровне моста находятся ядра черепно-мозговых нервов. Через него проходят нервные пути, соединяющие вышележащие отделы с продолговатым и спинным мозгом. Позади моста расположен мозжечок, с функцией которого в основном связывают координацию движений, поддержание позы и равновесия. Усиленный рост мозжечка отмечается на первом году жизни ребенка, что определяется формированием в течение этого периода дифференцированных и координированных движений. В дальнейшем темпы его развития снижаются. К 15 годам мозжечок достигает размеров взрослого. Средний мозг (мезенцефалон) включает ножки мозга, четверохолмие и ряд скоплений нервных клеток (ядер). В области четверохолмия расположены первичные центры зрения и слуха, осуществляющие локализацию источника внешнего стимула. Эти центры находятся под контролем вышележащих отделов мозга. Они играют важнейшую роль в раннем онтогенезе, обеспечивая первичные формы сенсорного внимания. Ядра (черная субстанция и красное ядро) играют важную роль в координации движений и регуляции мышечного тонуса. В среднем мозге расположена так называемая сетчатая, или ретикулярная, формация. В ее состав входят переключательные клетки, аккумулирующие информацию от афферентных путей. Восходящие пути от клеток ретикулярной формации идут во все отделы коры больших полушарий, оказывая тонические активирующие влияния. Это так называемая неспецифическая активирующая система мозга, которой принадлежит важная роль в регуляции уровня бодрствования, организации непроизвольного внимания и поведенческих реакций. Передний мозг состоит из промежуточного мозга (диэнцефалона) и больших полушарий. Промежуточный мозг включает две важнейшие структуры: таламус (зрительный бугор) и гипоталамус (подбугровая область). Гипоталамус играет важнейшую роль в регуляции вегетативной нервной системы. Вегетативные эффекты гипоталамуса, разных его отделов имеют неодинаковые направленность и биологическое значение. При функционировании задних отделов возникают эффекты симпатического типа, при функционировании передних отделов — эффекты парасимпатического типа. Восходящие влияния этих отделов также разнонаправлены: задние оказывают возбуждающее влияние на кору больших полушарий, передние — тормозящее. Связь гипоталамуса с одной из важнейших желез внутренней секреции — гипофизом — обеспечивает нервную регуляцию эндокринной функции. В клетках ядер переднего гипоталамуса вырабатывается нейросекрет, который по волокнам гипоталамо-гипофизарного пути транспортируется в нейрогипофиз. Этому способствуют и обильное кровоснабжение, и наличие сосудистых связей гипоталамуса и гипофиза. Гипоталамус принимает участие в регуляции температуры тела, водного обмена, обмена углеводов. Ядра гипоталамуса участвуют во многих сложных поведенческих реакциях (половые, пищевые, агрессивно-оборонительные). Гипоталамус играет важную роль в формировании основных биологических мотиваций (голод, жажда, половое влечение), а также положительных и отрицательных эмоций. Многообразие функций гипоталамуса дает основание расценивать его как высший подкорковый центр регуляции жизненно важных процессов, их интеграции в сложные системы, обеспечивающие целесообразное приспособительное поведение. Дифференцировка ядер гипоталамуса к моменту рождения не завершена и протекает в онтогенезе неравномерно. Развитие ядер гипоталамуса заканчивается в период полового созревания. Таламус составляет значительную часть промежуточного мозга. Это многоядерное образование, связанное двусторонними связями с корой больших полушарий. В его состав входят три группы ядер. Релейные ядра передают зрительную, слуховую, кожно-мышечно-суставную информацию в соответствующие проекционные области коры больших полушарий. Ассоциативные ядра связаны с деятельностью ассоциативных отделов коры больших полушарий. Неспецифические ядра (продолжение ретикулярной формации среднего мозга) оказывают активизирующее влияние на кору больших полушарий. Центростремительные импульсы от всех рецепторов организма (за исключением обонятельных), прежде чем достигнут коры головного мозга, поступают в ядра таламуса. Здесь поступившая информация перерабатывается, получает эмоциональную окраску и направляется в кору больших полушарий. К моменту рождения большая часть ядер зрительных бугров хорошо развита. После рождения размеры зрительных бугров увеличиваются за счет роста нервных клеток и развития нервных волокон. Онтогенетическая направленность развития структур промежуточного мозга состоит в увеличении их взаимосвязей с другими мозговыми образованиями, что создает условия для совершенствования координационной деятельности его различных отделов и мозга в целом. В развитии промежуточного мозга существенная роль принадлежит нисходящим влияниям коры больших полушарий. Базальные ганглии (хвостатое ядро, полосатое тело, бледный шар) играют важнейшую роль в осуществлении двигательной функции, являясь связующим звеном между ассоциативными и двигательными областями коры больших полушарий. Большие полушария головного мозга у взрослого человека составляют 80 \% массы головного мозга. Они соединены пучками нервных волокон, образующих мозолистое тело. В глубине больших полушарий расположена старая кора — гиппокамп, являющийся одной из важнейших структур лимбической системы. Лимбическая система, функционально объединяющая гиппокамп, гипоталамус, некоторые ядра таламуса и области коры, является важнейшей частью регуляторного контура (система структур, участвующих в регуляции нервных процессов в коре больших полушарий). Лимбическая система участвует в когнитивных, аффективных и мотивационных процессах. Основной структурой больших полушарий является новая кора (неокортекс), покрывающая их поверхность.
Кора больших полушарий
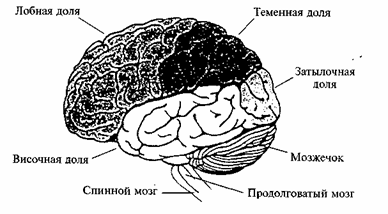
Кора больших полушарий представляет собой тонкий слой серого вещества на поверхности полушарий. В процессе эволюции поверхность коры интенсивно увеличивалась по размеру за счет появления борозд и извилин. Общая площадь поверхности коры у взрослого человека достигает 2200—2600 см2. Толщина коры в различных частях полушарий колеблется от 1,3 до 4,5 мм. В коре насчитывается от 12 до 18 млрд нервных клеток. Отростки этих клеток образуют огромное количество связей, что создает условия для обработки и хранения информации. В коре каждого из полушарий выделяют четыре доли — лобную, теменную, височную и затылочную (рис. 45). Каждая из этих долей содержит функционально различные корковые области.
Рис. 45. Доли коры
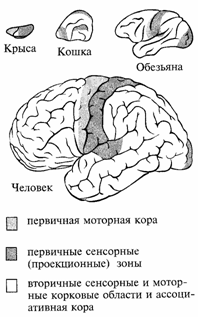
Проекционные сенсорные зоны, включающие первичные и вторичные корковые поля, принимают и обрабатывают информацию определенной модальности от органов чувств противоположной половины тела (корковые концы анализаторов по И. П. Павлову). К их числу относятся зрительная кора, расположенная в затылочной доле, слуховая — в височной, соматосенсорная — в теменной доле (рис. 46). Моторная кора каждого полушария, занимающая задние отделы лобной доли, осуществляет контроль и управление двигательными действиями противоположной стороны тела. Ассоциативные области составляют у человека основную часть поверхности коры больших полушарий (третичные поля). На рис. 47 видно, как нарастает их удельный вес в эволюционном ряду. Именно с этими областями связано формирование познавательной деятельности и психических функций. Клинические наблюдения показывают, что при поражении заднеассоциативных областей нарушаются сложные формы ориентации в пространстве, конструктивная деятельность, затрудняется выполнение всех интеллектуальных операций, которые осуществляются с участием пространственного анализа (счет, восприятие сложных смысловых изображений). Поражение лобных отделов коры приводит к невозможности осуществления сложных программ поведения, требующих выделения значимых сигналов на основе прошлого опыта и предвидения будущего. В ассоциативных областях коры левого полушария выделяются поля, непосредственно связанные с осуществлением речевых процессов, — центр Вернике в задневисочной коре, осуществляющий восприятие речевых сигналов, и центр Брока в нижних отделах лобной области коры, связанный с произнесением речи.
Рис. 46. Проекционные, ассоциативные и моторные области коры больших полушарий
Рис. 47. Изменение соотношения проекционных ассоциативных областей в эволюционном ряду Нейронная организация коры больших полушарий. В коре больших полушарий человека различные специализированные типы нейронов и их отростки пространственно организованы и распределены по шести слоям (рис. 48). I слой состоит в основном из конечных разветвлений апикальных дендритов пирамидных нейронов. Во II слое сосредоточено значительное количество вставочных клеток с разветвленной системой дендритов, связанных с пирамидными нейронами II и III слоя. Это некрупные афферентные пирамиды. В IV и V слое расположены пирамиды большого размера, коллекторы информации, посылающие эфферентные волокна другим нейронам. Наиболее крупные пирамиды находятся в V слое двигательной коры (гигантские клетки Беца). Их длинные аксоны формируют пирамидный тракт (VI слой), проводящий импульсы, по которым осуществляется управление движениями.
Рис. 48. Слои коры. Слева нейроны с отростками (окраска по Гольджи), в центре — тела нейронов разного типа и размера (окраска по Нисслю), справа — волокнистые структуры (окраска, выявляющая миелиновую оболочку)
Клетки разного типа, находящиеся в разных слоях коры, объединены большим количеством разнообразных связей и образуют определенные группировки — модули или ансамбли. В сенсорных проекционных отделах и моторной коре в объединениях преобладает вертикальная ориентация, определяемая апикальным дендритом. Это так называемые колонки или микроансамбли, в которых осуществляются аналитические процессы. Кроме микроансамблей выделены более сложные группировки (лестничные, гнездные), включающие большое количество нейронов разных типов, и разветвленные базальные дендриты (рис. 49). Такие ансамбли чаще встречаются в ассоциативных областях и являются структурной основой более сложной обработки информации.
Рис. 49. Нейронные группировки (ансамбли) в коре больших полушарий
Помимо внутриансамблевых межнейрональных связей, группировки нейронов имеют внешние связи. Выходящие за пределы ансамблей терминальные аксоны образуют системы ассоциативных связей, благодаря которым происходит объединение нейронных ансамблей как внутри одной корковой зоны, так и между зонами. Сложная разветвленная система внутрикорковых ассоциативных связей создает основу пластичной функциональной интеграции и системной организации деятельности мозга.
Созревание мозга в онтогенезе ребенка
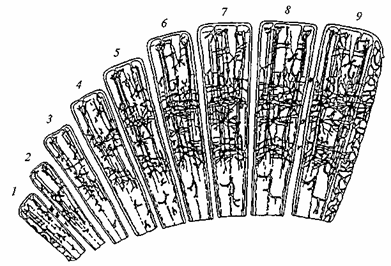
Головной мозг как многоуровневая структура неравномерно созревает в ходе индивидуального развития. Во внутриутробном периоде одновременно с закладкой и развитием основных жизненно важных органов первыми начинают формироваться отделы мозга, где расположены нервные центры, обеспечивающие их функционирование (продолговатый мозг, ядра среднего и промежуточного мозга). К концу внутриутробного периода у человека определенной степени зрелости достигают первичные проекционные поля. К моменту рождения уровень зрелости структур мозга позволяет осуществлять как жизненно важные функции (дыхание, сосание и др.), так и простейшие реакции на внешние воздействия, т.е. осуществляется принцип минимального и достаточного обеспечения функций. Закономерный ход созревания структур мозга в пренатальном периоде обеспечивает нормальное индивидуальное развитие, нарушения созревания приводят к ближайшим и отдаленным неблагоприятным последствиям, проявляющимся в нервно-психическом статусе и поведении ребенка. В постнатальном периоде продолжается интенсивное развитие мозга, в особенности его высших отделов — коры больших полушарий. Нейронная организация коры больших полушарий в онтогенезе. В развитии коры больших полушарий выделяются два процесса — рост коры и дифференцировка ее нервных элементов. Наиболее интенсивное увеличение ширины коры и ее слоев происходит на первом году жизни, постепенно замедляясь и прекращаясь в разные сроки — к 3 годам в проекционных, к 7 годам в ассоциативных областях. Рост коры происходит за счет увеличения межнейронального пространства (разрежение клеток) в результате развития волокнистого компонента (роста и разветвления дендритов и аксонов) и клеток глии, осуществляющей метаболическое обеспечение развивающихся нервных клеток, которые увеличиваются в размерах. Процесс дифференцировки нейронов, начинаясь также в раннем постнатальном периоде, продолжается в течение длительного периода индивидуального развития, подчиняясь как генетическому фактору, так и внешнесредовым воздействиям. Первыми созревают афферентные и эфферентные пирамиды нижних слоев коры, позже — расположенные в более поверхностных слоях. Постепенно дифференцируются различные типы вставочных нейронов. Раньше созревают веретенообразные клетки, переключающие афферентную импульсацию из подкорковых структур к развивающимся пирамидным нейронам. Звездчатые и корзинчатые клетки, обеспечивающие взаимодействие нейронов и циркуляцию возбуждения внутри коры, созревают позже. Заканчиваясь возбудительными и тормозными синапсами на телах нейронов, эти клетки создают возможность структурирования импульсной активности нейронов (чередование разрядов и пауз), что является основой нервного кода. Дифференцировка вставочных нейронов, начавшаяся в первые месяцы после рождения, наиболее интенсивно происходит в период от 3 до 6 лет. Их окончательная типизация в переднеассоциативных областях коры отмечается к 14-летнему возрасту. Функционально важным фактором формирования нейронной организации коры больших полушарий является развитие отростков нервных клеток — дендритов и аксонов, образующих волокнистую структуру. Аксоны, по которым в кору поступает афферентная импульсация, в течение первых трех месяцев жизни покрываются миелиновой оболочкой, что существенно ускоряет поступление информации к нервным клеткам проекционной коры. Вертикально ориентированные апикальные дендриты обеспечивают взаимодействие клеток разных слоев, и в проекционной коре они созревают в первые недели жизни, достигая к 6-месячному возрасту III слоя. Дорастая до поверхностных слоев, они образуют конечные разветвления. Базальные дендриты, объединяющие нейроны в пределах одного слоя, имеют множественные разветвления, на которых образуются множественные контакты аксонов других нейронов. С ростом базальных дендритов и их разветвлений увеличивается воспринимающая поверхность нервных клеток. Специализация нейронов в процессе их дифференциации и увеличение количества и разветвленности отростков создают условия для объединения нейронов разного типа в клеточные группировки — нейронные ансамбли. В нейронные ансамбли включаются также клетки глии и разветвления сосудов, обеспечивающие клеточный метаболизм внутри нейронного ансамбля. В развитии коры и формировании ансамблевой организации в онтогенезе выделяют следующие этапы (рис. 50). К моменту рождения вертикально расположенные пирамидные клетки в нижнем слое и их апикальные дендриты создают прообраз колонки, которая у новорожденных бедна межклеточными связями.
Рис. 50. Возрастные преобразования ансамблевой организации коры больших полушарий от рождения до 20 лет. Схема построена на основе результатов морфологического исследования мозга человека 1 — новорожденные; 2—3 мес. жизни; 3 — 6 мес.; 4 — 1 год; 5—3 года; 6 — 5—6 лет; 7— 9—10 лет; 8 — 12—14 лет; 9— 18—20 лет
1-й год жизни характеризуется увеличением размеров нервных клеток, дифференциацией звездчатых вставочных нейронов, увеличением дендритных и аксонных разветвлений. Выделяется ансамбль нейронов как структурная единица, окруженная тонкими сосудистыми разветвлениями. К 3 годам ансамблевая организация усложняется развитием гнездных группировок, включающих разные типы нейронов. В 5—6 лет наряду с продолжающейся дифференциацией и специализацией нервных клеток нарастают объем горизонтально расположенных волокон и плотность капиллярных сетей, окружающих ансамбль. Это способствует дальнейшему развитию межнейрональной интеграции в определенных областях коры. К 9—10 годам усложняется структура отростков интернейронов и пирамид, увеличивается разнообразие ансамблей, формируются широкие горизонтальные группировки, включающие и объединяющие вертикальные колонки. В 12—14 лет в нейронных ансамблях четко выражены разнообразные специализированные формы пирамидных нейронов, высокого уровня дифференцировки достигают интернейроны; в ансамблях всех областей коры, включая ассоциативные корковые зоны, за счет разветвлений отростков удельный объем волокон становится значительно больше удельного объема клеточных элементов. К 18 годам ансамблевая организация коры по своим характеристикам достигает уровня взрослого человека. Закономерности созревания структур мозга в онтогенезе. Основная закономерность в характере созревания мозга как многоуровневой иерархически организованной системы проявляется в том, что эволюционно более древние структуры созревают раньше. Это прослеживается в ходе созревания структур мозга по вертикали: от спинного мозга и стволовых образований головного мозга, обеспечивающих жизненно важные функции, к коре больших полушарий. По горизонтали развитие идет от проекционных отделов, включающихся в обеспечение элементарных контактов с внешним миром уже с момента рождения, к ассоциативным, ответственным за сложные формы психической деятельности. Для развития каждого последующего уровня необходимо полноценное созревание предыдущего. Так, для созревания проекционной коры необходимо формирование структур, через которые поступает сенсорно-специфическая информация. Для развития в онтогенезе ассоциативных корковых зон необходимо формирование и функционирование первичных проекционных отделов коры. Так, нарушение в раннем возрасте проекционных корковых зон приводит к недоразвитию областей более высокого уровня (вторичные проекционные и ассоциативные отделы). Этот принцип развития структур мозга в онтогенезе Л. С. Выготский обозначил как направление «снизу вверх». Следует подчеркнуть, что позже созревающие структуры не просто надстраиваются над уже существующими, а влияют на их дальнейшее развитие. Так, при исследовании активности отдельных нейронов было показано, что только после созревания проекционной корковой зоны нейроны релейного ядра таламуса приобретают специализированную реакцию зрелого типа в ответ на афферентный стимул. Сформированная многоуровневая организация мозга носит иерархический характер. Ведущую роль в осуществлении целостной интегративной функции мозга приобретают высшие отделы коры больших полушарий, управляющие подчиненными им структурами более низкого уровня. Такой принцип иерархической организации структур зрелого мозга Л.С. Выготский обозначил как направление «сверху вниз». Длительный и гетерохронный характер созревания структур мозга определяет специфику функционирования мозга в различных возрастных периодах.
Методики изучения функциональной организации мозга
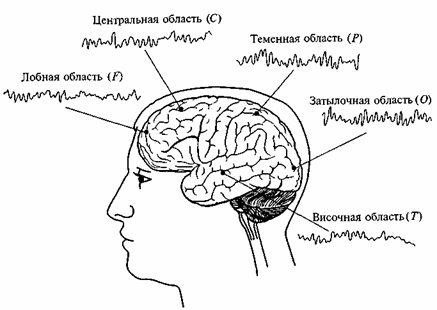
Одним из первых методов оценки функциональной роли разных структур мозга явился метод повреждения или удаления участков мозга с помощью хирургических, химических и температурных воздействий. Другой рано возникший метод — это метод прямой электрической стимуляции, который применялся как в экспериментах на животных, так и во время нейрохирургических операций, когда находящийся в сознании больной мог оценить свои ощущения при раздражении различных точек коры и подкорковых структур. Например, при раздражении проекционной зрительной коры больной как бы видел цветовые пятна, вспышки пламени; стимуляция вторичных зрительных полей вызывала сложные зрительные образы, раздражение определенных подкорковых ядер — звуковые и зрительные галлюцинации. С помощью электрической стимуляции во время операции была уточнена локализация речевых зон, физиологические основы речи, памяти и эмоций. Электроэнцефалография. В настоящее время наиболее распространенным и адекватным для изучения функциональной организации мозга является метод регистрации электроэнцефалограммы (ЭЭГ) — суммарной биоэлектрической активности, отводимой с поверхности головы. Многоканальная запись ЭЭГ в различных отведениях позволяет одновременно регистрировать электрическую активность функционально различных областей коры (рис. 51). В ЭЭГ выделяются следующие типы ритмических колебаний: дельта-ритм 0,5—3 Гц; тета-ритм 4—7 Гц; альфа-ритм 8—13 Гц, основной ритм ЭЭГ, преимущественно выраженный в каудальных отделах коры (затылочной и теменных); бета-ритм 15—30 Гц; гамма-колебания > 30—60 Гц. Эти ритмы различаются не только по своим частотным, но и функциональным характеристикам. Их амплитуда, топография, соотношение являются важным диагностическим признаком и критерием функциональной активности различных областей коры при реализации психической деятельности. Подробно этот вопрос будет рассмотрен в соответствующих главах. Анализ ЭЭГ осуществляется как визуально, так и с помощью ЭВМ. Визуальная оценка применяется в клинической практике. С целью унификации и объективизации диагностических оценок используется метод структурного анализа нативной ЭЭГ, основанный на выделении функционально сходных признаков и их объединении в блоки, отражающие характер активности структур мозга различных уровней (коры больших полушарий, диэнцефальных, лимбических, стволовых). В возрастной нейрофизиологии этот метод успешно используется для оценки степени структурно функциональной зрелости мозга.
Рис. 51. Электрическая активность, зарегистрированная от различных областей коры мозга человека. В скобках указаны латинские обозначения областей коры
В настоящее время как в клинических, так и в исследовательских целях широко используются компьютерные методы анализа ЭЭГ, позволяющие оценить выраженность различных ритмов по их спектральной мощности и их статистическую взаимосвязь (корреляционный анализ и анализ функции когерентности ритмической активности). Оценка когерентности ритмической активности широко используется в исследовательских целях. Этот метод позволяет выявить степень сходства организации ритмов биоэлектрической активности в различных мозговых структурах. Сходство организации рассматривается как необходимая предпосылка взаимодействия и адекватный показатель функциональной организации структур мозга при осуществлении различных видов деятельности. Рост значений функции когерентности (КОГ) биопотенциалов в ряде областей коры отражает увеличение вероятности их функциональной интеграции. Вызванные потенциалы. Другой тип суммарной электрической активности — вызванные потенциалы (ВП). Они возникают в ответ на внешние воздействия и отражают изменения функциональной активности областей коры, осуществляющих прием и обработку поступающей информации. ВП представляет собой последовательность разных по полярности — позитивных и негативных — компонентов, возникающих после предъявления стимула (рис. 52). Количественными характеристиками ВП являются латентный период (время от начала воздействия стимула до достижения максимального значения компонента) и амплитуда компонента. Компоненты ВП принято обозначать латинскими буквами по полярности: N— негативные, Р— позитивные, — и цифровыми индексами — по величине латентности в миллисекундах. Например, положительный компонент с латентным периодом 300 мс после предъявления стимула обозначается как Р300. Метод регистрации ВП широко используется при анализе процесса восприятия. В экспериментальных моделях на животных при одновременной регистрации ВП и активности отдельных нейронов была показана связь основного комплекса ВП с возбудительными и тормозными процессами, протекающими на разных уровнях коры больших полушарий. Было обнаружено, что начальные компоненты ВП — это так называемые экзогенные компоненты, связанные с активностью пирамидных клеток, которые воспринимают сенсорную информацию. Возникновение других, более поздних, фаз ответа обусловлено обработкой информации, осуществляемой нейронными аппаратами коры при участии не только сенсорного афферентного потока, но и импульсации, поступающей из других отделов мозга, в частности из ассоциативных и неспецифических ядер таламуса, и по внутрикорковым связям из других корковых зон.
50 мс Рис. 52. Зрительный вызванный потенциал. Начало ответа совпадает с моментом предъявления светового стимула
Эти нейрофизиологические исследования положили начало широкому использованию ВП человека для анализа процесса восприятия. У человека ВП имеет относительно небольшую амплитуду по сравнению с амплитудой фоновой ЭЭГ, и его изучение стало возможно только при использовании компьютерной техники для выделения сигнала из шума и последующего усреднения реакций, возникающих в ответ на ряд однотипных стимулов. ВП, регистрируемые при предъявлении сложных сенсорных сигналов и решении определенных когнитивных задач, получили название связанных с событиями потенциалов (ССП). При изучении ССП наряду с анализом параметров, используемых при анализе ВП, — латентного периода и амплитуды компонентов — используются и другие специальные методы обработки, позволяющие в сложной конструкции ВП выделить компоненты, связанные с определенными когнитивными операциями: метод главных компонент и метод разностных кривых. Метод главных компонент основан на использовании факторного анализа, позволяющего выделить факторы, наиболее тесно связанные с определенными операциями и приходящиеся на временной интервал, соответствующий тому или иному компоненту ССП. Это позволяет вычленить функциональную роль данного компонента в анализируемом процессе. С той же целью используется метод разностных кривых. Он заключается в следующем: сначала регистрируют фоновую кривую ССП при нейтральной стимуляции, а затем — кривую ССП при предъявлении конкретных задач. Потом с помощью компьютера эти две кривые сравнивают, и по преимущественной выраженности определенных компонентов делается заключение об их связи с выполняемой задачей. Топографическое картирование. Многоканальная регистрация ЭЭГ дает возможность представить полученные в результате компьютерной обработки ЭЭГ данные в удобном для восприятия виде — как карты одномоментного пространственного распределения по коре мощности разных ритмов ЭЭГ и амплитуд компонентов ВП или других характеристик. Последовательность таких карт дает представление о динамике процессов. На топографических картах мозга цветом и его интенсивностью кодируются различные параметры ЭЭГ. Такая визуализация позволяет охарактеризовать функциональную организацию мозга при разных состояниях и видах деятельности. Компьютерная томография основана на использовании вычислительной техники и новейших технических методов, позволяющих получить множество объемных изображений одной и той же структуры мозга. Из методов компьютерной томографии наиболее часто используется метод позитронно-эмиссионной томографии (ПЭТ). Этот метод позволяет охарактеризовать активность различных структур мозга на основе изменения метаболических процессов. При обменных процессах в нервных клетках используются определенные химические элементы, которые можно пометить радиоизотопами. Усиление активности сопровождается усилением обменных процессов, и в областях повышенной активности образуется скопление изотопов, по которым и судят об участии тех или иных структур в психических процессах (рис. 53). Другим широко используемым методом является ядерно-магнитно-резонансная томография. Метод основан на получении изображения, отражающего распределение плотности ядер водорода (протонов), при помощи электромагнитов, расположенных вокруг тела человека. Водород является одним из химических элементов, участвующих в метаболических процессах, и потому его распределение в структурах мозга — надежный показатель их активности. Преимущество ядерно-магнитно-резонансного метода состоит в том, что его использование не требует введения в организм радиоизотопов, и вместе с тем этот метод позволяет получить четкие изображения «срезов» мозга в различных плоскостях, так же как и метод ПЭТ. 36. Вегетативная нервная система это: ТолкованиеПеревод · · · · · · · · Вегетативная нервная система Вегетативная нервная система, которая также называетсяавтономной (systema nervosum autonomicum), контролирует растительные функции организма — такие, какпитание, дыхание, циркуляция жидкостей, выделение, размножение. Она иннервирует преимущественновнутренние органы и состоит из двух основных отделов: симпатического и парасимпатического. Совместнаяработа обоих отделов регулируется и контролируется корой головного мозга, которая является высшимотделом центральной нервной системы. Центры вегетативной нервной системы располагаются в головном испинном мозге. Выделяют также периферическую часть, которую составляют нервы, нервные окончания,сплетения и узлы. Центры симпатического отдела (pars sympathica) находятся в боковых рогах серого вещества грудного ипоясничного отделов спинного мозга. В передних корешках спинного мозга находятся преганглионарныесимпатические волокна, идущие от центров к предпозвоночным и околопозвоночным узлам симпатическогоотдела. Околопозвоночные узлы (ganglia trunci sympathici) проходят вдоль всего позвоночного столба, отуровня основания черепа до вершины копчиковой кости, и соединяются друг с другом межузловыми ветвями(rr. interganglionares), образуя два симпатических ствола (truncus sympathicus) (рис. 269) — левый и правый.В зависимости от локализации узлов в каждом симпатическом стволе выделяют несколько отделов. Шейный отдел симпатического ствола (pars cervicalis trunci sympathici) включает в себя верхний, средний инижний узлы, располагающиеся по обеим сторонам от позвоночного столба. Постганглионарные волокна,идущие от узлов, направляются по ходу артериальных ветвей головы, шеи и груди и образуют сплетения.Также постганглионарные волокна от каждого узла входят в состав верхнего, среднего и нижнегосимпатических нервов, которые направляются к сердцу и вместе с парасимпатическими и соматическиминервами образуют сердечные сплетения. Грудной отдел симпатического ствола (pars thoracica trunci sympathici) образован 10—12 симпатическимиузлами неправильной треугольной формы, располагающимися возле головок ребер. 1—5-й узлы даютпостганглионарные волокна, направляющиеся к грудной аорте и формирующие вокруг нее симпатическоесплетение. От этого сплетения, следуя вдоль ветвей грудной аорты, к пищеводу, бронхам и легкимнаправляются отдельные сплетения. Постганглионарные волокна 6—9-го узлов объединяются и образуютбольшой внутренностный нерв (n. splanchnicus major) (рис. 268). Волокна 10—12-го узлов образуют малыйвнутренностный нерв (n. splanchnicus minor) (рис. 268). Внутренностные нервы через щели диафрагмыпроникают в брюшную полость и принимают участие в образовании чревного сплетения. Брюшной, или поясничный, отдел симпатического ствола (pars abdominalis trunci sympathici) состоит изпоясничных симпатических узлов, располагающихся по четыре с каждой стороны. Они залегают на переднейповерхности тел поясничных позвонков и соединяются между собой поперечными и продольнымимежузловыми ветвями. Постганглионарные волокна двух верхних поясничных узлов принимают участие вобразовании чревного, или солнечного, сплетения (plexus coeliacus). Помимо этих волокон, в составсолнечного сплетения входят внутренностные нервы, постганглионарные волокна 1—2-го поясничныхсимпатических узлов, а также волокна парасимпатического ядра блуждающего нерва. От солнечногосплетения вдоль ветвей чревной и верхней брыжеечной артерий направляются одноименные сплетения,иннервирующие поджелудочную железу и кишечник до нисходящей ободочной кишки. А вдоль парных ветвейбрюшной артерии сплетения направляются к почкам, надпочечникам и половым железам. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|