|
|
Б. Метаболические функцииМитохондрии являются «силовой станцией» клетки, поскольку за счет окислительной деградации питательных веществ в них синтезируется большая часть необходимого клетке АТФ (АТР). В митохондриях локализованы следующие метаболические процессы: превращение пирувата в ацетил-КоА, катализируемое пируватдегидрогеназным комплексом: цитратный цикл; дыхательная цепь, сопряженная с синтезом АТФ (сочетание этих процессов носит название «окислительное фосфорилирование»); расщепление жирных кислот путем β-окисления и частично цикл мочевины. Митохондрии также поставляют клетке продукты промежуточного метаболизма и действуют наряду с ЭР как депо ионов кальция, которое с помощью ионных насосов поддерживает концентрацию Са2+в цитоплазме на постоянном низком уровне (ниже 1 мкмоль/л). Главной функцией митохондрий является захват богатых энергией субстратов (жирные кислоты, пируват, углеродный скелет аминокислот) из цитоплазмы и их окислительное расщепление с образованием СО2 и Н2О, сопряженное с синтезом АТФ. Реакции цитратного цикла приводят к полному окислению углеродсодержащих соединений (СО2) и образованию восстановительных эквивалентов, главным образом в виде восстановленных коферментов. Большинство этих процессов протекают в матриксе. Ферменты дыхательной цепи, которые реокисляют восстановленные коферменты, локализованы во внутренней мембране митохондрий. В качестве доноров электронов для восстановления кислорода и образования воды используются НАДН и связанный с ферментом ФАДН2. Эта высоко экзергоническая реакция является многоступенчатой и сопряжена с переносом протонов (Н+) через внутреннюю мембрану из матрикса в межмембранное пространство. В результате на внутренней мембране создаетсяэлектрохимический градиент. В митохондриях электрохимический градиент используется для синтеза АТФ из АДФ (ADP) и неорганического фосфата (Рi) при катализе АТФ-синтазой.
Способность образовывать многочисленные выпячивания является характерной чертой митохондрий. Такие выпячивания ( кристы ) чаще всего имеют вид плоских гребней. Кристы участвуют в преобразовании энергии питательных вещ-в, поступающих в клетку, в энергию молекул.
Эндоплазматическая сеть — это обязательная органелла эукариотической клетки. Она обнаружена в клетках растений, животных и человека. Функции этой составляющей части клетки разнообразны и связаны в основном с синтезом, модификацией и транспортом органических соединений. Впервые эндоплазматическая сеть была обнаружена в 1945 году. Американский ученый К. Портер разглядел ее с помощью одного из первых электрических микроскопов. С этого времени началось ее активное исследование. В клетке есть две разновидности этой органеллы: Гранулярная, или шероховатая эндоплазматическая сеть (покрыта множеством рибосом). Агранулярная, или гладкая эндоплазматическая сеть. Каждый тип ретикулума имеет некоторые особенности и выполняет совершенно разные функции. Давайте рассмотрим их более подробно. Невероятная польза бананов для здоровья Как поднять самооценку: 12 простых шагов Ответ на главный вопрос: почему вы не можете похудеть Гранулярная эндоплазматическая сеть: строение. Данная органелла представляет собой системы цистерн, пузырьков и канальцев. Стенки ее состоят из билипидной мембраны. Ширина полости может колебаться от 20 нм до нескольких микрометров — здесь все зависит от секреторной активности клетки. У мало специализированных клеток, которые характеризируются низким уровнем метаболизма, ЭПС представлена всего лишь несколькими разрозненными цистернами. Внутри клетки, которая активно синтезирует белок, эндоплазматическая сеть состоит из множества цистерн и разветвленной системы канальцев. Как правило, гранулярная ЭПС посредством канальцев связана с мембранами ядерной оболочки — именно таким образом происходят сложные процессы синтеза и транспорта белковых молекул. Гранулярная эндоплазматическая сеть: функции. Как уже упоминалось, вся поверхность ЭПС со стороны цитоплазмы покрыта рибосомами, которые, как известно, участвуют в синтезе белка. ЭПС — это место синтеза и транспорта протеиновых соединений. Что может случится с мозгом из-за мобильного телефона Что в течение часа делает с организмом баночка "Кока-Колы"? О чем больше всего сожалеют люди в конце жизни Эта органелла отвечает за синтез интегральных белков цитоплазматической мембраны. Но в большинстве случаев созданные белковые молекулы далее с помощью мембранных пузырьков транспортируются в комплекс Гольджи, где происходит их дальнейшая модификация и распределение соответственно потребностям клетки и тканей. Кроме того, в полостях цистерн ЭПС происходят и некоторые изменения белка — например, присоединение к нему углеводного компонента. Здесь же, путем агрегации образуются большие секреторные гранулы.
В электронном микроскопе плазмолемма выглядит как типичная биологическая мембрана, состоящая из двух электронноплотных слоев, между которыми находится электроннопрозрачный слой. Общая толщина всех трех слоев в плазмолемме составляет 12-14 нм. Однако надо отметить, что эта трехслойная структура является лишь основой плазмолеммы, поскольку к ней снаружи и изнутри примыкают слабоконтрастируемые молекулярные комплексы. Плазмолемма обладает асимметрией, которая выражается в различиях состава липидов и белков наружной и внутренней частей. Асимметрия плазмолеммы проявляется у животных клеток, в частности, в том, что они имеют на своей внешней поверхности особый углеводный слой – гликокаликс. Он образован входящими в состав мембраны олигосахаридами и липидами в комплексе с белками. Гликокаликс присутствует у всех животных клеток, однако степень его развития может быть разной. В наибольшей степени он развит у всасывающих клеток кишечного эпителия. Гликокаликс этих клеток создает среду для пристеночного пищеварения, а также защищает плазмолемму от повреждений. Вирус гриппа содержит фермент нейраминидазу, который необходим для удаления одного из компонентов гликокаликса - сиаловой кислоты, препятствующей прикреплению вирусных частиц к плазмолемме эпителиальных клеток. Гликокаликс обладает выраженными антигенными свойствами, что облегчает опознание клеток при их взаимодействии между собой.
С внутренней стороны плазмолемма связана с такими компонентами цитоскелета, как микротрубочки и микрофиламенты. Это позволяет животной клетке не только поддерживать определенную форму, но и активно изменять ее. Взаимодействие плазмолеммы с цитоскелетом лежит в основе активного движения фибробластов и макрофагов, удлинения проходящих по капиллярам эритроцитов, изменения формы клетки при фагоцитозе и секреции. Плазмолемма, отграничивая содержимое клетки от внешней среды, одновременно обеспечивает избирательный обмен веществ между клеткой и средой. Транспорт веществ через плазмолемму осуществляется при помощи различных механизмов.
Бокаловидные клетки — один из энтероцитов эпителия кишечника. Располагаются на кишечных ворсинках поодиночке среди каёмчатых энтероцитов. Обычно отсутствуют на самой вершине ворсинки, а также на дне крипты. В тонкой кишке человека составляют 9,5 % от всех клеток эпителиального пласта. Число бокаловидных клеток увеличивается в дистальном направлении кишки.[3] Кроме кишечника, бокаловидные клетки имеются в слизистой оболочке дыхательных путей, в конъюнктиве глаз, протоках поджелудочной и околоушных слюнныхжелёз.[4] В дыхательных путях больше всего бокаловидных клеток находится в экстраторакальной части трахеи, с уменьшением диаметра бронхов их количество существенно сокращается, а вбронхиолах менее 1 мм их нет совсем.[5] Структура и функции Бокаловидные клетки накапливают гранулы муциногена, которые, абсорбируя воду, набухают и превращаются в муцин (основной компонент слизи). При этом клетки обретают форму бокала, суженного у основания (где находится ядро) и округлой широкой в апикальной, верхней части. Затем набухшая верхняя часть бокаловидной клетки разрушается, слизь переходит в просвет органа, клетка приобретает призматическую форму и снова начинает накапливать муциноген. Слизь, выделяемая бокаловидными клетками, служит для увлажнения поверхности слизистой оболочки кишечника и этим способствует продвижению химуса, а также участвует в процессах пристеночного пищеварения. Апикальная часть бокаловидных клеток имеет исчерченную каёмку, подобно каёмчатым энтероцитам. Однако, в отличие от каёмчатых энтероцитов, микроворсинки каёмки у бокаловидных клеток расположены реже и неодинаковы по высоте.
Набор хромосом в ядре соматической клетки, как патологический, так и нормальный называется кариотипом. Ученые из Швеции А. Леван и Дж. Тио ещё в 1956 году доказали, что во всех клетках человека, кроме половых, число хромосом равно 46. Этот хромосомный набор называется парным или диплоидным. Хромосомы человека подразделяются на половые и аутосомные. Аутосомные хромосомы соединены в пары, поскольку они содержат гены, которые отвечают за одинаковые функции. У человека их 22 пары или 44, а последняя пара – это половые хромосомы. В половой клетке человека хромосом в два раза меньше – их 23, то есть клетки являются гаплоидными, но при оплодотворении диплоидность клетки восстанавливается. Половые хромосомы могут быть двух типов: X и Y. Женская половая клетка имеет две Х хромосомы, а мужская – одну X и одну Y. Во всех парах хромосом, как половых, так и аутосомных одна из хромосом получена от матери, а другая – от отца. Следовательно, половая принадлежность ребенка зависит от того какой тип половой клетки мужчины (Y или Х) соединится с женской (Х). Строение хромосомы отчетливо видно во время клеточного деления, когда она сильно спирализована. Внешний вид хромосом существенно изменяется на разных этапах клеточного цикла. В хромосоме выделяют длинное (обозначается буквой «q» и короткое плечо (обозначается буквой «р»), концы которых называются теломерами. Было установлено, что теломеры крайне важны при сохранении стабильности хромосом, так как препятствуют слипанию их концов после редупликации. В фазе митоза хромосомы представлены парой сестринских хроматид, которые соединены центромерой. Она имеет весьма сложное строение и определяет движение хромосомы при делении. В центромере имеется особое вещество – кинетохор. Оно участвует в формировании нитей веретена при делении клетки. В зависимости от положения центромеры хромосомы подразделяются на несколько групп: метацентрические (центромера находится посередине хромосомы), акроцентрические (центромера находится ближе к концу хромосомы) и субметацентрические. Впервые деление кариотипа человека на группы проведено было на конференции в Денвере в 1960 году. В результате аномалии числа либо структуры хромосом человека возникают патологические состояния, которые называют хромосомными синдромами, которые не лечатся. Описано и изучено более трехсот хромосомных аномалий. Клинические проявления при аномалиях хромосом наблюдаются с рождения. Частота хромосомных аномалий на 1000 новорожденных составляет от 5-7%. Но этот показатель мог быть значительно выше, если бы каждый эмбрион с перестройками в хромосомах смог проходить полный цикл внутриутробного развития и появиться на свет. Хромосомные аномалии возникают не только в половых, но и в соматических клетках. Анализ хромосомного набора позволяет обнаружить нарушения, которые могут привести к аномалии развития, наследственным заболеваниям и даже гибели эмбриона и плода на ранних стадиях развития. Примерно 50% всех самопроизвольных абортов обусловлены хромосомной перестройкой у эмбриона, которая нарушает скоординированную работу генов в раннем эмбриогенезе. Поэтому около 30% оплодотворенных яйцеклеток гибнет в первые 10 дней после процесса оплодотворения. В первом триместре беременности 50% всех случаев выкидыша связана с аномалиями хромосом, а во втором триместре беременности - 25% случаев. Люди с аномалиями хромосом могут быть как весьма тяжелыми инвалидами, так и вполне полноценными членами общества. Все зависит от вида аномалии, общего состояния организма и заботы близких. Самой распространенной хромосомной аномалией считается синдром Дауна, который обусловлен наличием дополнительной хромосомы в клетках человека по причине трисомии по 21-ой паре. К весьма распространенным хромосомным синдромам относится также синдром Клайнфельтера (аномальный набор половых хромосом ХХУ у мальчиков) , синдром Патау (трисомия по 13-ой паре), Синдром Шерешевского-Тернера (женщины лишены одной хромосомы Х).
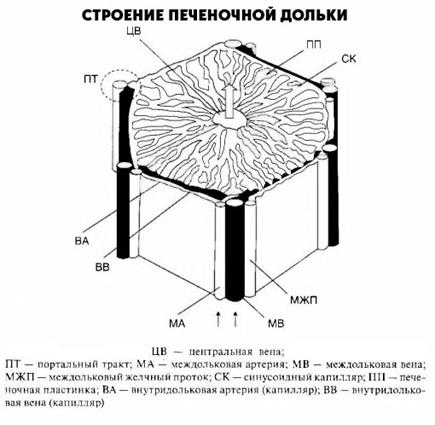
Печеночная долька - морфофункциональная единица печени. В центре дольки находится центральная вена. Центральные вены, соединяясь между собой, в итоге впадают в печеночные вены, последние, в свою очередь, впадают в нижнюю полую вену. Долька имеет форму призмы 1-2 мм. Она состоит из радиально расположенных двойных рядов клеток (печеночных пластин, или балок). Между рядами гепатоцитов находятся внутридольковые желчные протоки, их концы, обращенные к центральной вене, замкнуты. Образовавшаяся желчь направляется к периферии долек. Между печеночными пластинами находятся синусоидные капилляры, где смешивается кровь, поступающая в печень по воротной вене и собственной печеночной артерии. По периферии печеночной дольки находятся триады: междольковые вены (до которых разветвляется воротная вена), междольковые артерии (до которых разветвляется собственная артерия печени) и междольковые желчные протоки (которые, сливаясь между собой, в итоге формируют правый и левый печеночные протоки). Таким образом, внутри печеночной дольки желчь продвигается от центра к периферии и в дальнейшем через общий желчный проток выводится из печени. Кровь из воротной вены и собственной артерии печени, смешиваясь внутрипеченочной дольки, двигается от ее периферии к центру и выводится через центральные вены в систему нижней полой вены. Печеночная долька отграничена от других соединительнотканной оболочкой, содержащей коллагеновые и эластиновые волокна. Общее число печеночных долек составляет около 0,5 млн. За 1 мин через печень взрослого протекает 1,2 л крови, почти 70% из которой поступает через воротную вену.
Для клинической оценки важным является состояние синусоидов. Они имеют три отдела: периферический, промежуточный и центральный. Промежуточный отдел составляет 90% их длины. Он, в отличие от периферического и центрального отдела, не имеет базальной мембраны. Между эндотелием синусоида и гепатоцитами имеются пространства, сообщающиеся с пери портальными пространствами; вместе с межклеточными щелями они служат началом лимфатической системы. Именно в указанных пространствах и совершается контакт различных веществ с цитоплазматической мембраной печеночной клетки. Эндотелий синусоидов содержит поры, обеспечивающие переход в гепатоциты различных молекул. Некоторые из эндотелиальных клеток обеспечивают структуру синусоидов, а другие, например звездчатые ретикулоэндотелиоциты (клетки Купфера), обладают фагоцитарной функцией или принимают участие в обновлении и новообразовании соединительной ткани. Указанные клетки составляют 40% всех эндотелиальных клеток. При этом 48% эндотелиальных клеток выполняют структурную функцию и 12% - фибропластическую. Периферические отделы печеночной дольки образованы мелкими гепатоцитами, они участвуют в процессе регенерации и исполняют роль пограничной пластинки, отделяя паренхиму дольки от соединительной ткани портального поля. Через пограничную пластинку в дольку проникают междольковые вены системы v. portae и артериолы печеночной артерии, выходят холангиолы, впадающие в междольковые печеночные протоки. Между гепатоцитами и соединительной тканью находятся пространства, именуемые пространствами Моля. Портальный тракт на периферии дольки имеет вид треугольника с заключенными в нем конечными ветвями портальной вены, печеночной артерии и междольковым желчным протоком, называемыми триадой. В ее состав входят лимфатические щели с выстланным эндотелием и нервы, оплетающие кровеносные сосуды. Богатая сеть нервных волокон проникает в печеночные дольки до гепатоцитов и эндотелиальных клеток. Соединительная ткань в виде ретикулиновых и коллагеновых волокон, а также базальных мембран синусоидов, кровеносных сосудов и желчных протоков портального тракта у детей очень нежная и лишь у пожилых людей образует грубые волокнистые скопления.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
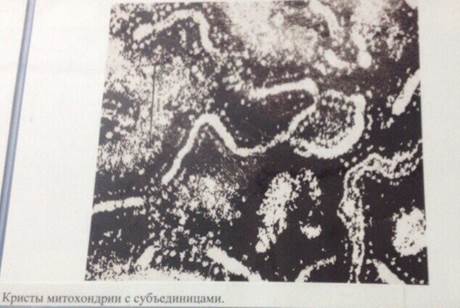 Внутренняя мембрана митохондрий образует выросты в полость в виде пластин или трубок, называемых кристами. Кристы бывают различных типов. Пространство между кристами заполнено однородным прозрачным веществом - матриксом митохондрий.
Внутренняя мембрана митохондрий образует выросты в полость в виде пластин или трубок, называемых кристами. Кристы бывают различных типов. Пространство между кристами заполнено однородным прозрачным веществом - матриксом митохондрий.
 Плазмолемма (цитолемма, плазматическая мембрана) создает селективный барьер между клеткой и внешней средой. Уже в начале XIX в. было обнаружено набухание и сжатие клеток, погруженных в растворы различной ионной силы, что свидетельствовало о наличии на поверхности клетки полупроницаемой мембраны. В 30-е гг. XX в. было доказано, что она состоит из липидов и белков, причем липиды образуют в ней бимолекулярный слой. Плазмолемма непроницаема для макромолекул, поэтому белки цитоплазмы создают в клетке осмотическое давление, под действием которого вода непрерывно поступала бы в клетку, если бы вне клетки не поддерживалась уравновешивающая концентрация других веществ. Это равновесие создается, прежде всего, молекулярным насосом, который выкачивает из клетки ионы натрия и закачивает в клетку ионы калия. З счет разности концентраций ионов внутри и вне клетки плазмолемма приобретает потенциал до +85 мВ.
Плазмолемма (цитолемма, плазматическая мембрана) создает селективный барьер между клеткой и внешней средой. Уже в начале XIX в. было обнаружено набухание и сжатие клеток, погруженных в растворы различной ионной силы, что свидетельствовало о наличии на поверхности клетки полупроницаемой мембраны. В 30-е гг. XX в. было доказано, что она состоит из липидов и белков, причем липиды образуют в ней бимолекулярный слой. Плазмолемма непроницаема для макромолекул, поэтому белки цитоплазмы создают в клетке осмотическое давление, под действием которого вода непрерывно поступала бы в клетку, если бы вне клетки не поддерживалась уравновешивающая концентрация других веществ. Это равновесие создается, прежде всего, молекулярным насосом, который выкачивает из клетки ионы натрия и закачивает в клетку ионы калия. З счет разности концентраций ионов внутри и вне клетки плазмолемма приобретает потенциал до +85 мВ. Бокаловидные клетки (синонимы: энтероциты бокаловидные, бокаловидные экзокриноциты; лат. enterocytus caliciformis) — продуцирующие слизь клетки эпителия слизистой оболочки кишечника и других органов позвоночных животных и человека[1]. Бокаловидные клетки часто называют одноклеточными железами[2].
Бокаловидные клетки (синонимы: энтероциты бокаловидные, бокаловидные экзокриноциты; лат. enterocytus caliciformis) — продуцирующие слизь клетки эпителия слизистой оболочки кишечника и других органов позвоночных животных и человека[1]. Бокаловидные клетки часто называют одноклеточными железами[2]. Локализация
Локализация акустических (слуховых) сигналов.
б) Как следует из приведённого названия клетки,
акустических (слуховых) сигналов.
б) Как следует из приведённого названия клетки,

 Хромосома – это структурная составляющая ядра клетки, которая содержит дезоксирибонуклеиновую кислоту (ДНК) – носитель наследственной информации, а также белок и немного РНК.
Хромосома – это структурная составляющая ядра клетки, которая содержит дезоксирибонуклеиновую кислоту (ДНК) – носитель наследственной информации, а также белок и немного РНК.
 Функциональная единица включает в себя синусоид с окружающим пространством между его эндотелием и гепатоцитами (пространство Диссе), прилежащими гепатоцитами и желчным канальцем. Некоторые авторы полагают, что структуру печени следует рассматривать исходя из строения приводящих и отводящих кровеносных сосудов, их переплетения,
Функциональная единица включает в себя синусоид с окружающим пространством между его эндотелием и гепатоцитами (пространство Диссе), прилежащими гепатоцитами и желчным канальцем. Некоторые авторы полагают, что структуру печени следует рассматривать исходя из строения приводящих и отводящих кровеносных сосудов, их переплетения,