|
Глава 9. МОРФОФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ СИСТЕМЫ ПИЩЕВАРЕНИЯ РЫБ
Филогенетическое развитие отдельных групп рыб проходило в резко различающихся условиях среды. Часть рыб оказалась и морской, часть - в пресной воде. Рыбы освоили водоемы всех географических и климатических зон на Земле с температурами атмосферного воздуха от минус 50 до плюс 50 њС. Естественно, формирование разнообразных водоемов сопровождалось параллельным формированием специфических биоценозов, т. е. кормовых баз, к которым рыбы были вынуждены приспосабливаться. Это, в свою очередь, привело к возникновению морфофункционального разнообразия рыб и прежде всего их систем пищеварения.
Кроме своей основной функции пищеварительный тракт рыб участвует в процессе газообмена, осморегуляции, размножения, защитных реакциях и др.
§39. ОБЩАЯ ХАРАКТЕРИСТИКА ПИЩЕВАРИТЕЛЬНОГО ТРАКТА РЫБ
Строение пищеварительного тракта у рыб проще, чем у высших позвоночных. Тем не менее внутри класса рыб имеется огромное разнообразие видовых особенностей, довольно сложных в морфологическом отношении. Из-за этого огромного разнообразия морфологии до сих пор невозможно признать, что строение и функции желудочно-кишечного тракта рыб изучены хорошо. В; обобщенном виде пищеварительная система рыб представлена на рис. 9.1.
Понимая, что разнообразие морфологических особенностей в пищеварительном тракте рыб требует систематизации, исследователи предлагают ряд классификаций. Одной из наиболее удачных представляется классификация Г. Г. Вундш, к которой специалисты обращаются с 1937 г. В соответствии с ней все известные виды рыб имеют один из 5 типов пищеварительной системы;
1. Лососевый тип (стенка желудка тонкая; имеется от 80 до 400 пилорических придатков).
2. Окуневый тип (толстостенная глотка; цилиндрический желудок; имеется только 3 пилорических придатка).
3. Щуковый тип (толстостенный пищевод; удлиненный желудок; печень вытянута в соответствии с геометрией тела).
4. Карповый тип (пищеварительный тракт имеет вид тонкой трубки, которая образует несколько петель; желудка нет, но передний отдел кишки расширен).
5. Угревый тип (узкий мускульный пищевод окружен печенью).
Гистологически пищеварительная трубка рыб не отличается от таковой других позвоночных. Пищевод состоит из слизистого, подслизистого, мышечного и серозного слоев. Эпителиальный слой включает в себя клетки реснитчатого эпителия, большое количество мукозных клеток. Здесь же обнаруживаются вкусовые рецепторы, кубические и грушевидные клетки.
Задний отдел пищевода (эзогастер) у некоторых видов имеет секреторные железы желудочного типа. В слизистой встречаются и разветвленные трубчатые железы.
Пищевод для большинства видов рыб- транзитный участок пищеварительной трубки. Однако у некоторых видов он выполняет добавочные функции: респираторную, депонирующую, а также первичной переработки пищи.
По типу пищеварения рыб делят на две группы - желудочные (преимущественно хищные) и безжелудочные (рис. 9.2).
И те и другие встречаются в пределах одного семейства и даже одного рода. Морфология желудка и присутствие в нем пищеварительных желез не всегда связаны с природой питания рыбы. Эпителий желудка описан подробно. Он имеет простое строение, состоит из цилиндрических клеток, обладает щеточной каймой, включает в себя мукозные клетки.
В желудочных железах костистых рыб идентифицирован только один тип секреторных клеток, которые продуцируют и зимогены, и соляную кислоту. На апикальном конце клеток, образующих железы, имеется компактная система канальцев, сходных со структурами обкладочных клеток у высших животных, вырабатывающих соляную кислоту. В базальной масти этих же клеток обнаружены гранулы, содержимое которых высвобождается в апикальной части клетки. Гистохимический анализ под-твердил предположение о том, что у рыб пепсиноген и соляную кислоту вырабатывают одни и те же клетки желудочных желез.

Рис. 9.1. Топография органов пищеварения у костистой рыбы:
1-аорта; 2- 4- сердце; 5- печень; 6- желудок; 7- тонкая кишка; 8- селезенка; 9- пилорические придатки; 10- гонады, 11 - мочевой пузырь; 12 - плавательный пузырь, 13 - почка; 14- желчный пузырь; 15-17- клоака; 18-жабры; 19- кювьеров проток Рис. 9.2. Разновидности пищеварительного тракта у разных видов рыб:
а- скат; б - осетр; в - лосось: г - окунь; д- кари; е - толстолобик; 1 - спиральный клапан; 2- пилорическая железа; 3- пилорические придатки
Кишечник у примитивных рыб - простейшая трубка, площадь которой увеличивается за счет спирального клапана - продольной складки слизистой оболочки. Количество оборотов спирали может быть от 2 до 50.
У костистых высокоорганизованных видов площадь кишки увеличивается за счет ряда приспособ-лений:
пилорических придатков у желудочных рыб; количество придатков колеблется от 1 до 1000; их слизистая не проявляет ферментативной активности (рис. 9.3);
продольных петель кишки;
складок слизистой оболочки.
Длина кишки у хищных и всеядных рыб примерно равна длине тела. У бентософагов и макрофитофагов длина кишки может превышать длину тела в 2-3 раза. Известно и 15-кратное превышение. При голодании рыб длина кишки уменьшается на 30-45 %.
Гистологи в кишечной стенке рыб выделяют 4 слоя. Однако подслизистый слой, например, у карпа может и не обнаруживаться. Мышечный слой лучше развит у хищных видов. Слизистая кишечника выстлана однослойным цилиндрическим эпителием с реснитчатой каймой. Помимо энтероцитов в составе слизистой описаны бокаловидные, грушевидные и кубические клетки, которые лишены щеточной каймы, а также клетки-мигранты-лимфоциты и различные гранулоциты. Для многих видов типичны клетки реснитчатого эпителия, присутствие которого считается признаком примитивной организации.
Кишечные железы у большинства рыб отсутствуют.
Терминальная часть кишечника называется прямой или задней кишкой. У некоторых видов этот участок кишки отделен особым клапаном. У реснитчатого окуня имеется даже гладкомышечный сфинктер, разделяющий среднюю кишку на две части с разным гистологическим строением.

Рис. 9.3. Топография пилорических придатков
У костистых рыб задняя кишка имеет слизистую, сходную по гистостроению. Энтероциты кишечника рыб принято делить на зимогенные и абсорбирующие. Есть сообщения о третьем типе энтероцитов - клетках с ярко выраженными эндокринными свойствами.
Застенные пищеварительные железы рыб представлены только печенью и поджелудочной железой. Эти органы ввиду топографической близости называют гепатопанкреасом.
Печень рыб развивается из эпителия пищеварительной трубки. Чаше она разделена на доли. Гепатоциты рыб имеют сходное строение с клетками печени других позвоночных. При химическом анализе в них обнаруживается большая концентрация липидов и гликогена, Большинство видов рыб имеют желчный пузырь, проток которого открывается в переднюю часть кишечника или (при их наличии) в пилорические придатки. Сюда же открываются и мелкие протоки поджелудочной железы. Сама поджелудочная железа заполняет пространства между петлями кишки вблизи печеночных долей и селезенки. Эндокринная часть поджелудочной железы у рыб имеет свои границы в отличие от островкового строения ее у высших позвоночных.
§40. ВЛИЯНИЕ ХАРАКТЕРА ПИТАНИЯ НА МОРФОЛОГИЮ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
Эволюционное влияние пищевого спектра на строение желудочно-кишечного тракта у рыб проявляется на уровне ротового аппарата и головной кишки. У круглоротых сравнительно простые челюсти образованы передними висцеральными дугами. У костистых рыб они имеют более сложное строение. Механическая переработка пищи в ротовой полости у рыб встречается в редких случаях. Движение челюстей обеспечивают кусание, разгрызание, выброс изо рта инородных тел. а также нагнетание воды в ротовую полость с целью прокачивания ее через жабры. Строение рта отражает особенности питания (рис. 9.4),
Так, у бентософагов рот нижний (осетр), у планктонофагов- верхний (пелядь).
Характер питания отразился и на строении туловищной кишки. Так, наиболее характерным признаком большинства хищных рыб является наличие хорошо развитого мускульного желудка и связанного с ним пепсиногенного пищеварения. Однако многие хищные рыбы утратили желудок. Существует несколько гипотез, объясняющих исчезновение этого органа. Одни ученые считают причиной исчезновения желудка переход рыб на питание моллюсками с известковой раковиной и, следовательно, нейтрализацию кислого желудочного сока.

Рис. 9.4. Расположение рта на голове у разных видов рыб:
а- верхний; б- конечный; в - полунижний; г - нижний; д- выдвижной; е -круглый рот
Другие ученые видят причину утраты желудка рыб в высоком проценте балласта (ил, песок, детрит) в пище.
Наконец, у всеядных рыб при частой перемене спектра питания и необходимости в более длительном пребывании пищи в кишке кислотный характер пищеварения (желудок) становится бесполезным
§41. ФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
Состав секретируемых желудочно-кишечным трактом соков сложен и ввиду ряда технических и методических причин до конца не изучен у рыб, как и механизм самой секреции. Секреторная интенсивность отмечена и в головной, и в туловищной кишке рыб. Однако интенсивность секреции и химическая реактивность секретируемых продуктов в разных участках пищеварительной системы, конечно, неодинаковы.
Немногие виды рыб имеют механизм секреции в ротовой полости, глотке и пищеводе. У большинства рыб в ротовой полости обнаруживается секрет, представленный только слизью. Главное назначение такого секрета - защита эпителия и вкусовых рецепторов.
Чем грубее пища, составляющая обычный рацион рыбы, тем более развит секреторный аппарат. Так, у рыб, питающихся кораллами (скаровые рыбы), отмечается интенсивная секреция слизи в ротовой полости. Обильная слизь способствует проглатыванию кораллов.
Обильное слизевыделение характерно и для цихлид, однако слизь у них играет несколько иную роль. Она необходима для выкармливания молоди. Лабиринтовые рыбы используют такую же ротовую слизь для построения гнезда. Показатель рН ротовой слизи рыб близок к нейтральной величине.
Слизистая оболочка пищевода имеет множественные борозды и складки, в которых накапливается слизь, задача которой заключается в обеспечении движения пищевого кома из ротовой полости после проглатывания.
У некоторых видов рыб (кефаль) слизистая пищевода имеет секреторные клетки, аналогичные желудочным секреторных клеткам.
Секреторная активность желудка. Секреторная функция желудка у рыб существенно отличается от таковой у наземных теплокровных животных.
Хотя эта функция желудка и находится под нейрогуморальным контролем, как и у других животных, у рыб имеется своя специфика, вызванная их таксономическим положением и образом жизни. Растяжение стенки желудка приводит к возбуждению пищеварительных центров в центральной нервной системе, По блуждающему нерву к секреторному аппарату желудка поступает стимул.
В регуляции секреции не меньшую роль играет и местная эндокринная система желудка, которая посредством биологически активных веществ - гастрина, гистамина, ацетилхолина - стимулирует секрецию желудочного сока.
Ацетилхолин является первым исполнителем воли центра. Он обладает разносторонним действием. Прежде всего он выступает в роли медиатора в синапсах. Ацетилхолин способен непосредственно возбуждать секреторные клетки желудка. Он стимулирует выделение гастрина, вызывает усиление синтеза гистамина.
Тастрин в 1,5 раза активнее гистамина стимулирует выделение соляной кислоты. Оба слабо влияют на отделение пепсиногена. Гастрин действует на клетки слизистой желудка через кровь, т. е. с определенной временной задержкой.
Тистамин обладает паракриновым эффектом в отношении секретирующих клеток желудка, т.е. действует непосредственно и, следовательно, быстро.
Основными компонентами желудочного сока рыб являются слизь, ферменты и соляная кислота. Физиологическая роль соляной кислоты исключительна и сводится к следующему.
Соляная кислота активирует зимогены и превращает, например, пепсиноген в пепсин - активную форму.
Соляная кислота создает оптимальное значение рН в желудке, что важно для максимально эффективной работы ферментов. По данным разных исследователей, рН желудочного сока колеблется от 1,2 до 5,0. Пепсин максимально активен при рН 1,0-2,0.
Соляная кислота способствует набуханию пищи, растворяет кости, кальцинированные кожные образования - чешую, жучки, а также наружный скелет и панцирь моллюсков, кораллов и т. д., а также участвует в регуляции процесса эвакуации желудочного содержимого в кишечник.
Однако анализ пищеварения хищников из семейства карповых (голавль, жерех), у которых нет желудка и не вырабатывается соляная кислота, позволяет говорить о том, что она не является обязательным компонентом пищеварительных соков даже у хищных рыб.
У некоторых рыб, например у акул, концентрация соляной кислоты в желудочном соке достигает 3 %. Поэтому тушка жертвы вынутая из желудка акулы через несколько минут после проглатывания, уже лишена чешуи, жучек и других кальцинированных образований.
Количество секретируемого пепсина у рыб зависит от температуры воды. Как резкое повышение, так и понижение температуры воды уменьшают секрецию фермента.
Количество соляной кислоты и слизи в большей мере определяется количеством поступившей в желудок пищи. Механическое растяжение желудка сопровождается усилением секреции соляной кислоты.
Помимо протеолитических ферментов в желудке рыб обнаружена липаза. В желудке некоторых видов, например угря, радужной форели, судака, ставриды, обнаруживают фермент хитиназу. Однако нет доказательств ее эндогенного происхождения. Хитиназная активность желудка - скорее результат автолиза. Как известно, автолиз широко распространен в природе.
Еще одна особенность желудочного пищеварения рыб - высокая лизосомная активность, что для рыб, питающихся зоопланктоном с высоким содержанием различных полисахаридов в качестве строительного материала покровных тканей, очень важно.
Секреторная функция кишечника. Секреторная функция кишечника рыб изучена недостаточно. Пищеварительные соки, обнаруживаемые в кишке, есть смесь секретов самой кишки, поджелудочной железы и печени. Дополнительную сложность при изучении кишечной секреции создают пилорические придатки, у которых имеется собственный секреторный аппарат.
Известно, что секрецию кишечника реализуют энтероциты и слизистые клетки кишечного эпителия, причем первые вырабатывают энзимогены, вторые - слизь. Считается, что энтероциты рыб продуцируют ферменты всех трех групп: протеолитические, липолитические, гликолитические.
Основным протеолитическим ферментом кишечного сока является трипсин. Поскольку, как и лю-бой другой фермент, он вырабатывается в форме профермента, то у него должен быть активатор. Таковым в кишечном соке является другая протеаза - энтерокиназа. В свою очередь, активный трипсин выполняет роль активатора по отношению к другим зимогенам кишечного сока и поджелудочной железы, например хемотрипсиногену,
В опытах in vitro слизистая кишечника проявляет и липолитическую активность, что позволяет говорить о секреции энтероцитами липаз. Липазы обнаружены и в пилорических придатках.
Гликолитические ферменты кишечного сока представлены рядом ферментов, проявляющих активность по отношению к разным углеводам. Причем этот спектр богаче у растительноядных рыб. Так, кишечник карпа секретирует амилазу, глюкозидазу, мальтазу, сахаразу, лактазу. Некоторые авторы относят сюда и целлобиазу, что вызывает сомнение.
Амилаза обнаружена в кишечном соке хищных, например форели, что объясняет успешное выращивание этого вида на кормах, содержащих углеводы.
Хитиназа обычна в кишечном соке карповых. Однако действует она специфично, не до конца. В экскрементах карпа обнаруживают оболочки яиц артемий, эфиппиумы дафний, челюсти хирономид. Скорее всего, хитиназа (???) обеспечивает расчленение наружного скелета насекомых и их личинок. Речь об использовании хитина как питательного вещества вести нельзя. А потому возникает вопрос: хитиназа ли это вообще?
Кроме отмеченных выше, вероятно, кишечник рыб секретирует щелочную фосфатазу и нуклеазы. Однако действие этих ферментов у рыб не изучено.
В регуляции кишечной секреции большую роль играют гормоны и электролиты. В опытах денервация кишечника не влияла на секрецию в кишечнике. Однако степень наполнения кишки влияет на секрецию посредством возбуждения Мейснеровского сплетения, расположенного в подслизистом слое. Следовательно, регуляция кишечной секреции имеет все ту же нейрогуморальную природу.
Конечная (прямая) кишка имеет слизистую, образованную эпителием, продуцирующим слизь.
У безжелудочных рыб кишечная секреция непрерывна, у желудочных - периодична, связана с эвакуацией химуса из желудка.
Характеристика ферментных систем. Гидролиз белков. У желудочных рыб гидролиз белков начинается с денатурации белковых молекул соляной кислотой с последующим расщеплением кислыми карбоксильными протеазами. В желудке основной протеазой является пепсин, который вырабатывается клетками слизистой в неактивной форме, т. е. в виде зимогена.
Под влиянием ионов водорода или протеаз в результате ограниченного протеолиза при рН 2,0-5,0 происходит превращение пепсиногена в пепсин. Активация зимогена связана с отщеплением от N-конца 44 аминокислотных остатков, что приводит к образованию пепсина, после "информационных изменений молекулы которого образуется активный центр. Существует несколько молекулярных форм пепсина (например, у трески - 3, у кошачьей акулы - 4). Молекулярная масса зимогена 40 000, активированного фермен-та 35000. Пепсин в желудке гидролизует около 10% всех пептидных связей. Для рыб характерна активность ферментов в широком спектре рН(1,0-7,0) и температур (0-30 њС).
В содержимом желудка обнаруживаются и другие ферменты, но механизм их секреции у рыб не описан. Достоверно известно, что у хищных (акул) большую роль играет фермент коллагеназа.
Более глубокому перевариванию белков способствуют ферменты поджелудочной железы и кишечного сока.
У безжелудочных рыб схема переваривания несколько иная: она не имеет стадий воздействия кислых протеаз. Из ротовой полости Пища поступает сразу в нейтральную или слабощелочную среду туловищ-ной кишки. Наиболее сильному воздействию химус подвергается со стороны панкреатических ферментов - трипсина, хемотрипсинов А, В и С, эластазы, а также карбоксипептидаз Аи В
Подобно желудочным пептидазам протеолитические ферменты поджелудочной железы выделяются а виде зимогенов, которые путем ограниченного протеолиза при помощи каскадного механизма превращаются в активные формы.
Зимоген трипсиноген активируется кишечной энтерокиназой и трипсином, остальные протеазы - только трипсином.
Молекулярная масса трипсина рыб от 21 400 (карп) до 26000 (амурский сом). Трипсин карпа активен при рН 4,7 и 9,2. Активность трипсина карпа в 3-7 раз выше таковой млекопитающих.
Трипсин расщепляет связи в основном между основными аминокислотами, хемотрипсин - связи ароматических аминокислот, эластаза -связи между алифатическими аминокислотами. Несмотря на то что аминокислотный состав энзимов довольно разнообразен (трипсиноген - 229 аминокислотных остатков, хемотрипсиноген - А - 245, эластаза - 240), ключевую роль в катализе играют 3 аминокислотных остатка, входящих в состав активного центра, - серина, гистидина и аспарагина.
Указанные ферменты, кроме того, обладают амилазной и эстеразной активностью.
Нативные белки плохо расщепляются трипсином и хемотрипсином. Они становятся доступными для гидролиза только после денатурации за счет изменения вторичной и третичной структур.
Зимоген прокарбопепсидаза А поджелудочной железы имеет 2 формы: с молекулярной массой 87 000 и 64 000.
Как желудочные, так и панкреатические пептидазы дробят молекулу белка до олигопептидов, состоящих из 2-6 аминокислотных остатков.
Пептидазы кишечного сока завершают процесс деполимеризации белковых молекул.
В кишечном соке содержится несколько пептидаз. Наиболее значимый фермент кишки - амино-пептидаза М (аланин-амино-пептидаза). Этот фермент отщепляет основные кислоты от олигопептидов.
Аминопептидазы А, имеющие молекулярную массу от 112000 до 350000, отщепляют остаток аспарагиновой и глутаминовой кислот.
Есть в кишечном соке и дипептидазы и энтерокиназы, имеющие свои особые функции.
Гидролиз углеводов. Оптимальным значением активной реакции среды для гидролитических про-цессов является рН от 6,5 до 7,5.
Субстратами для гликолитических ферментов кишечника и поджелудочной железы потенциально являются 3 группы веществ-
1. Структурные полисахариды (целлюлоза, лигнин, агар, хитин). Однако, согласно современным представлениям, эта группа углеводов не чувствительна к ферментам позвоночных. Если они деполимеризуются, то исключительно под влиянием энзимов, вырабатываемых микробами.
2. Универсальные полисахариды (крахмал, гликоген).
3. Олиго-, дисахариды и моносахара обладают способностью проходить через слизистую кишки без существенных изменений.
Гликолитические ферменты активны при рН, близком к 7, или в слабощелочной среде. Поэтому у желудочных рыб переваривание углеводов начинается только при поступлении корма в кишечник.
Тонкий механизм воздействия ферментов на углеводную цепочку у рыб не описан. Однако если признать, что общие принципы деполимеризации молекулы полисахаридов у высших и низших животных одни и те же, то следует предположить наличие в панкреатическом соке а- и у-амилазы.
Экспериментальные данные о происхождении ос-амилазы в кишечнике рыб противоречивы или видоспецифичны. Так, например, у щуки, судака отмечено наличие ос-амилазы в мембране кишки; у карпа, окуня, плотвы - в полости кишки. Как известно, а-амилаза является эндогидролазой, а у-амилаза - экзогидролазой. Причем а-амилаза разрывает полимер в средней части, действуя на 1,4-а-гликозидные связи с образованием больших обломков молекул. у-амилаза последовательно отщепляет остатки глюкозы от конечной части полимерного остатка после обработки а-амилазой.
У высших животных, имеющих в составе слюны или в рубце а-амилазу, в кишечник поступают олигосахариды, освобожденные а-амилазой, а также олигосахариды, содержащиеся в пище.
Кишечный и панкреатический соки из желудочно-кишечного тракта рыб содержат помимо у-амилазы мальтазу, сахаразу, лактазу, которые гидролизуют олигосахара до мономеров. Скорее всего, а-амилазная активность у рыб с желудком проявляется в пилорических придатках, а у безжелудочных рыб - раньше - в самом начале туловищной кишки.
Карбогидразы, осуществляющие заключительные этапы гидролиза углеводов, связаны с апикальной мембраной энтероцитов. Они синтезируются в цитоплазме энтероцитов в зоне базальной мембраны, затем транспортируются к апикальной мембране и встраиваются в нее в виде гликопротеиновых комплексов. Они имеют довольно большую молекулярную массу (около 200000) и существуют в виде макромолекулярных комплексов: мальтаза - у-амилаза, мальтаза - сахараза, сахараза - изомальтаза.
Гидролиз жиров. Пищевые жиры представлены чаще всего триглицеридами животного и растительного происхождения.
Начало гидролизу молекул триглицеридов дает панкреатическая липаза в полости кишки, в результате чего образуются моноглицерид и жирные кислоты. Процесс зависит от наличия желчных кислот (см. схему).
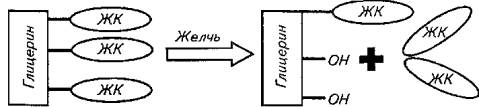
Панкреатическая фосфолипаза, активируемая трипсином, гидролизует эфирную связь глицерина и жирных кислот с образованием лецитина, изолецитина и жирных кислот.
Окончательное переваривание 2-моноглицерида осуществляют ферменты апикальной мембраны энтеропитов - моноглицерид-липаза, эстераза и др.
§42. МЕХАНИЗМ ПИЩЕВАРЕНИЯ
По механизму действия ферментов на субстрат, локализации энзимов в пищеварительной трубке, отношению пищеварения к клеточным мембранам и транспортным системам различают три типа пищеварения.
1. Внеклеточное пищеварение (полостное, дистантное) (впервые описано Spallanzani в 1783 г.).
2. Внутриклеточное пищеварение (подробно изучено И. И. Мечниковым).
3. Мембранное пищеварение (открыто А. М. У голевым в 1958г.).
Внеклеточное пищеварение. Пищеварение протекает во внеклеточной среде - в специальных полостях. За счет этого процесса происходит начальный этап деструктурирования биополимеров при помощи эндогидролаз.
Основными особенностями дистантного пищеварения являются наличие водных растворов, произвольная ориентация активных центров ферментов к субстрату, вероятностный характер распределения ферментов в полостях,
Полостное пищеварение эффективно по отношению к крупным молекулам. Однако его эффективность по отношению к олигомерам низкая. Другим недостатком внеклеточного пищеварения является разобщенность продуктов гидролиза и транспортных систем.
Следует, однако, подчеркнуть, что полостное пищеварение связано с мембранным. Эти два процесса не являются разобщенными. Однако их взаимоотношения изучены недостаточно.
Внутриклеточное пищеварение. Оно подразделяется на два типа-
Первый тип пищеварения реализуется за счет транспортирования небольших молекул через клеточные мембраны и последующего гидролиза ферментами цитоплазмы (например, пептидов).
Второй тип пищеварения связан со специализированными вакуолями клеток. Вакуоли могут постоянно присутствовать в составе протоплазмы, а иногда иметь временный характер, как, например, при фагоцитозе, пиноцитозе или микропиноцитозе. Подробнее эти явления будут рассмотрены ниже (см. §43).
Внутриклеточное пищеварение лимитировано низкой проницаемостью мембран энтероцитов. Согласно исследеваниям ученых, на ранних стадиях онтогенеза, например в личиночной стадии развития, внутриклеточное пищеварение является основным способом ассимиляции питательных веществ рыб.
Мембранное (контактное) пищеварение. Оно осуществляется в основном ферментами, активными к олигополимерам и находящимися на поверхности микроворсинок (рис. 9.5).
 
Рис. 9.5. Схема соотношения полостного и мембранного пищеварения:
а - пустой кишечник, б-кишечнике пищевыми частицами; I - область полостного пищеварения; II-область мембранного пищеварения; 1-полостные ферменты; 2- микроворсинки; 3- ферменты щеточной каймы; 4-поры щеточной каймы; 5- микробная клетка; 6 и 7- пищевые частицы
В мембрану микроворсинок, имеющую липидную природу, погружены белки и гликопротеиды, причем ее углеводная часть (гликокалекс) довольно сильно развита.
Мукополисахаридные нити соединены между собой кальциевыми мостиками, которые, периодически разрушаясь, способствуют проникновению крупных молекул в глубь гликокалекса.
В зоне гликокалекса выявлена высокая протеолитическая, липолитическая и гликолитическая активность, причем ферменты мембраны имеют строгую ориентацию - активным центром в полость. Часть ферментов, фиксированных на мембране, имеют особую "ножку" и как бы удалены от нее.
Преимущества мембранного (контактного) пищеварения следующие.
Гликокалекс защищает мембрану от воздействия химуса.
Ферменты мембран всегда ориентированы активным центром к субстрату.
Микроворсинки препятствуют проникновению в энтероцит клеток микробов и чужеродных белков.
При мембранном (контактном) пищеварении обеспечивается немедленное транспортирование продуктов гидролиза.
Щеточная кайма обеспечивает стабильные условия для активного функционирования фиксированных на ней ферментов (рН, химический и газовый составы водной среды, температура).
Соотношение мембранного и полостного пищеварения (по активности ферментов) различно у разных видов рыб. Оно также изменяется по сезонам и зависит от степени накормленности рыбы.
Как правило, соотношение мембранного (Мб) и полостного (Пл) пищеварения выше у хищных рыб. Например, у налима, щуки, судака и окуня соотношение Мб: Пл в течение года более единицы, у растительноядных и всеядных менее единицы.
Следовательно, мембранное пищеварение для хищников имеет большее значение, что не лишено биологического смысла. У хищных рыб кишечник короче и желудочно-кишечный тракт в целом имеет меньшие объем и площадь всасывания.
У хищников период активного питания наблюдается и зимой, т. е. пищеварение протекает в широких температурных границах, например в средней полосе от 0 до 30 њС. У хищных рыб высокая пищевая активность сопровождается интенсивными гидролитическими процессами, так как ферменты гликокалекса у них активны в широком диапазоне температур и рН.
Для сравнения укажем, что у таких мирных рыб, как карп, карась, плотва, при понижении температуры воды до 12 њС снижается пищевая активность на фоне резкой заторможенности пищеварительных процессов.
§43. СИМБИОТИЧЕСКОЕ ПИЩЕВАРЕНИЕ
Роль симбиотического пищеварения хорошо изучена у жвачных. Последние исследования показали, что оно не менее важную роль играет и у низших животных.
Общее количество микроорганизмов в пищеварительной системе рыб колеблется в широких пределах в течение года. Пик численности приходится на лето. Большое влияние оказывает и состав пищи. Так, например, в 1 г содержимого кишечника линя и белого амура содержится 1,29х1012 клеток, причем в естественной пище микробов больше, чем в комбикормах.
Роль микрофлоры в процессах пищеварения рыб возрастает по мере развития онтогенеза. На примере карпа, белого амура, линя и карася установлена родовая принадлежность микробов кишки: Pseudomonas, Micrococcus, Bacillus, Bacterium, Pseudobacterium, Azotobacter, Sarcine.
Доминируют представители рода Pseudomonas. Этот род связан с протеолитической активностью пищевого кома. Присутствие микрофлоры в желудочно-кишечном тракте объясняет и целлюлозолитическую и хитиназную активность химуса.
Микробная масса сама по себе является полноценным белком. Так, в состав микробного белка из кишки карпа входит 13 аминокислот, причем продукция свободных аминокислот микробного происхождения максимальна у сеголетков, минимальна у трехлетков карпа.
Микрофлора рыб способна к фиксации молекулярного азота. При интенсивном питании толстолобика установлена самая высокая нитрогеназная активность по всей длине кишки - 3,648 мг белка в час. Азотфиксация видоспецифична. Она изменяется по возрастающей: щука >карп, лещ, плотва >растительноядные.
Велика роль микрофлоры в синтезе витаминов. Около 50 % потребности рыб в витаминах обеспечивает микробный синтез,
§44. ВСАСЫВАНИЕ ПРОДУКТОВ ПИЩЕВАРЕНИЯ
Системы транспортирования продуктов пищеварения сходны у всех животных. Известно два типа транспортирования: макромолекулярный и микромолекулярный.
Первый тип обеспечивает перенос крупных молекул и надмолекулярных агрегаций через слой эпителиоцитов по межклеточным каналам. Главными механизмами макромолекулярного переноса служат фагоцитоз и пиноцитоз.
Фагоцитоз - это захват клеткой корпускулярных структур при помощи временных цитоплазматических выростов (псевдоподий) и воздействие на них цитоплазматических гидролитических ферментов. Фагоцитировать могут лейкоциты, энтероциты щеточной каймы. Фагоцитированию подвергаются лишь очень мелкие частицы (не более 1 мкм).
Пиноцитоз - это захват (обтекание) клеткой капелек жидкости с последующим их гидролизом. Экспериментально доказано, что у рыб все энтероциты способны к инвагинации и захвату жидкого содержимого. Благодаря пиноцитируюшей способности энтероциты рыб захватывают нативные или частично переваренные белковые молекулы. Помимо белков энтероциты рыб пиноцитируют и мельчайшие капельки жира. При помощи маркеров доказано пиноцитирование большого количества эмульсии, образующейся из жиров рациона рыбы и желчи, в состав которой входят липиды, липоиды, фосфолипиды, стерины и эфиры, а также продукты их неполного расщепления, например ди- и моноглицериды.
Топография пиноцитоза разнообразна. Состав пиноцитирующих пузырьков переднего отдела кишки представлен в основном жировыми веществами, а в задних отделах кишечника пиноцитозу в большей мере подвержены белковые структуры.
Основой транспортирования продуктов гидролиза является микромолекулярный тип, обеспечивающий перенос мономеров и олигомеров с небольшой молекулярной массой. Его обслуживают три физиологических механизма: пассивное транспортирование, облегченная диффузия, активное транспортирование.
Пассивное транспортирование объединяет процессы диффузии и осмоса. Реализуемое благодаря наличию градиента концентрации, электрохимическому градиенту и пор в мембране, оно протекает без затрат энергии. Скорость пассивного переноса обратно пропорциональна размеру молекул переносимого вещества. Чем мельче молекула, тем быстрее она проникает через клеточную мембрану. Таким способом транспортируются вода и минеральные вещества.
Облегченная диффузия осуществляется по градиенту электрохимического потенциала под контролем ферментативных систем. Прохождение диффундируемого вещества через клеточную мембрану облегчается другим веществом или молекулой. В качестве примера облегченной диффузии можно привести транспортирование глюкозы из крови в клетку. Процесс протекает против градиента концентрации. Транспортирование глюкозы облегчается транспортными белками клеточной мембраны.
Активное транспортирование требует затрат энергии. Оно осуществляется при помощи специальных транспортных систем всегда против градиента концентрации и электрохимического градиента. В кишечных клетках существуют транспортеры многих типов. Они переносят одну или несколько органических молекул сразу.
Мобильный переносчик представляет собой, как правило, высокомолекулярный белок. Источником энергии для транспортеров служит АТФ у всех животных независимо от организации. В редких случаях используется энергия градиента концентрации ионов (при транспортировании аминокислот и глюкозы).
Усвоение пищи у рыб при благоприятных условиях (оптимальная температура, химический и газовый составы воды, рН) достигает 80%. Переедание понижает этот показатель, недоедание, наоборот, повышает. При скудном питании у хищных отмечают высокую усвояемость даже белков костей и чешуи.
Растительная пища в целом менее доступна для организма рыб по сравнению с животной. А при высокой пищевой активности и избыточном потреблении пищи, например, у фитопланктонофагов в фекалиях обнаруживают жизнеспособные водоросли, т.е. степень усвоения пиши становится крайне низкой. Такую же картину наблюдают и с бактериально-протозойной фракцией планктонофагов.
У высших животных слизистая кишечника специализирована на транспортирование определенных групп веществ. У рыб выделяется лишь концевой участок туловищной кишки, в котором ограничено всасы-вание органических веществ и преобладает всасывание минеральных веществ и воды, хотя в целом в прямой кишке рыб химус обезвоживается не так интенсивно, как у высших животных. Влажность экскрементов у рыб составляет около 90 %.
Не нашли, что искали? Воспользуйтесь поиском по сайту:
©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|

