|
Изменение остроты зрения у гуппи с возрастом
| Длина рыбы, мм
| Разрешающий угол глаза, мин
| |
|
| |
|
| |
|
| |
|
| |
|
|
Сетчатка глаза у рыб устроена примерно так же, как и у высших позвоночных (рис. 2.8). Она имеет инвертированный характер восприятия света. Прежде чем попасть на светочувствительные клетки, расположенные в базальной части ретины, свет проходит через ганглионарные, биполярные и частично через амокринные и горизонтальные клетки в составе сетчатки. Несомненно, что при этом происходит частичное рассеивание света, Биологический смысл данного явления остается неясным, Однако ни один исследователь не берет на себя смелость заявлять об инвертированности сетчатки глаза как эволюционной ошибке природы.
Инвертированность сетчатки отмечается в строении глаза всех позвоночных животных от рыб до высших млекопитающих. Если бы инвертированность сетчатки была ошибкой природы, то в процессе эволюционирования от низших животных к высшим она была бы устранена за ненадобностью.
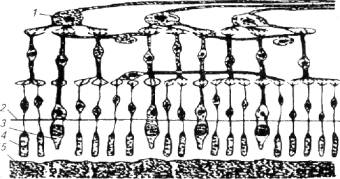
Рис. 2.8. Схема строения сетчатки глаза рыб:
ганглионарная клетка; 2- наружная пограничная мембрана;3- колбочка; 4- палочка; 5- пигментный эпителий
Как видно из рисунка 2.8, ретина имеет довольно сложное микростроение, представленное, по крайней мере, четырьмя функционально важными слоями специфических клеток (слой ганглионарных, биполярных, амокринных и горизонтальных клеток) осле проникновения светового луча через ганглионарные биамокринные и горизонтальные клетки квант света воспринимается слоем пигментного эпителия, клетки которого имеют включения красителя фусцина (реже гуанина). Пигментные клетки обладают подвижностью и могут подниматься или опускаться в соседний слой - слой рецепторных клеток, открывая или прикрывая их от световых лучей, т.е. слой пигментного эпителия служит фильтром (подобно темным солнцезащитным очкам у человека), дозирующим общую освещенность фоторецепторов.
Фоторецепторный слой клетчатки образован клетками трех типов палочками, одиночными колбочками и сдвоенными (близнецовыми) колбочками (Рис 2.9).
По мнению ряда авторов одиночные колбочки костистых рыб морфологически неоднородны и делятся на клетки с коротким миоидом и клетки с удлиненным миоидом
По сетчатке палочки и колбочки рассредоточены неравномерно. На периферии обнаруживаются только палочки. В центральной части сетчатки могут присутствовать и палочки, и колбочки. В области центральной ямки светочувствительный слой образован одними колбочками. Экспериментально установлено, что палочки обеспечивают восприятие световой энергии в условиях недостаточной освещенности (при сумеречном свете).
При высокой освещенности в работу включаются колбочки, обеспечивая при этом высокую остроту зрения и цветовое видение объектов. Соотношение палочек и колбочек в ретине неодинаково у разных рыб и определяется двумя факторами: эволюционным положением вида и образом жизни. Например, у многих пластинчатожаберных рыб ретина состоит исключительно из палочек.
 
Рис. 2.9. Фоторецепторные клетки рыб:
а - палочки; б- колбочки (одиночные и близнецовые); 1- наружный сегмент: 2- эллиисоид; 3- миоид (сократительный элемент); 4-наружная пограничная мембрана сетчатки 5-клеточное ядро; 6- масляная капля, содержащая каротиноид; 7- основание (ножка клетки)
У костистых морских глубоководных рыб количество колбочек очень небольшое. Если быть точным, то надо отметить и влияние стадии онтогенеза на соотношение светочувствительных клеток в сетчатке глаза рыб. На личиночной стадии развития у большинства рыб превалируют колбочки, а у некоторых, например сельдей, палочки вообще отсутствуют. И лишь при переходе на активное питание происходит разрастание палочек. Это биологически оправдано, так как у активно питающейся молоди из-за высокого уровня обменных процессов возникает потребность в пищевом поведении и в условиях ограниченной видимости.
Слой сетчатки глаза, образованный биполярными клетками, обеспечивает самый первый уровень интеграции сигналов, поступающих от фоточувствительных клеток - палочек и колбочек.
У активных дневных рыб с хорошим зрением четыре светочувствительных клетки образуют синапсы с четырьмя биполярными клетками, которые, в свою очередь, контактируют с одной ганглиозной клеткой.
У сумеречных рыб каждая биполярная клетка интегрирует большее количество фоторецепторов. Так, у налима на теле одной биполярной клетки обнаруживается 3-4 десятка синапсов с фоторецепторами, а на каждую ганглиозную клетку приходится 7 биполярных клеток.
Из вышесказанного ясно, что слой ганглиозных клеток сетчатки выполняет функцию дальнейшей интеграции зрительных сигналов. Аксоны ганглиозных клеток дают начало зрительному нерву, идущему в зрительные центры головного мозга.
Ретиномоторная реакция. Сетчатке рыб свойственна ретиномоторная реакция, позволяющая зрительному анализатору формировать адекватную зрительную картину независимо от степени освещенности наблюдаемого объекта. Этот механизм особенно важен Для рыб, так как возможности зрачка глаза как регулятора светового потока у рыб очень ограничены. Поэтому только ретиномоторная реакция дает рыбам возможность быть активными в условиях меняющегося освещения.
Схема адаптации ретины к уровню светового потока представлена на рис. 2.10.
При попадании на ретину яркого света после темновой адаптации (правая часть рис. 2.10) в сетчатке наблюдаются перемещения светочувствительных клеток. Первыми реагируют клетки пигментного эпителия: они погружаются в слой палочек и колбочек и закрывают палочки. При этом колбочки за счет своих сократительных структур - миоидов - подтягиваются и как бы вылезают из пигментного слоя. Палочки производят перемещения в противоположном от колбочек направлении, что обеспечивает их плотное укрытие пигментом. Таким образом, колбочки поглощают энергию яркого света, а палочки реципроктно блокируются и не участвуют в процессе фоторецепции.

Рис. 2.10. Ретиномоторная реакция:
1-палочки; 2- колбочки; 3- пигментные 1 клетки
В условиях ограниченной освещенности (левая часть рис. 2.10) к наружной мембране подтягиваются палочки, а колбочки погружаются в пигментный слой и изолируются от световых лучей.
Границы освещенности, при которой запускается ретиномоторный механизм, различны у разных рыб. У морской атерины ретиномоторная реакция возникает при освещенности 1-10лк, у ставриды - 0,01-1,0лк, у карася - 0,01 -1,0 лк, у вьюна - 0,01-0,1 лк, у голавля - 0,001-0,0001 лк.
На чувствительность глаза рыбы к яркости света влияет процесс темновой (световой) адаптации. Темновая адаптация глаза рыб занимает около 30 мин, световая адаптация происходит быстрее - всего за 10-30с. Однако полная настройка глаза на эффективную рецепцию света может продолжаться и несколько часов. Контрастная чувствительность глаза в сумерках ниже, чем при ярком освещении. Способность выделять предмет из общего] фона зависит от яркости фона, характеристик самого предмета (его яркости, размеров, подвижности) и адаптационного состояния глаза.
Выше уже отмечалось, что рыбы видят недалеко, если проводить сравнение с наземными животными. Здесь отметим следующее. Дальность видимости предметов в воде зависит и от их цветности. Сети, окрашенные в разный цвет, хамса, например, замечает на разном расстоянии (в м):
сине-зеленые 0.5-0,7
темно-синие 0,8-1,2
темно-коричневые 1,3 - 1,5
серые и черные 1,5-2,0
белые 2,0-2,5
Предметы в воде и предметы над водой воспринимаются рыбой неодинаково из-за разной преломляющей способности воды и воздушной среды. На рис. 2.11 показано поле зрения рыбы.
Предметы у поверхности воды рыба воспринимает крупнее, чем они есть на самом деле, а дальние объекты - как мелкие. Такая информация, несмотря на ее необъективность, не лишена смысла, так как объекты у поверхности воды представляют большую опасность для рыбы, чем удаленные объекты.
В поле зрения рыбы попадают предметы, даже расположенные у горизонта (дерево). Однако они воспринимаются рыбами в сильно искаженном виде. Зрительное поле рыбы настолько специфично, что в оптике широкоугольные объективы, создающие по периферии искажение, получили название "рыбий глаз".
В воде рыба (имеются в виду активные дневные виды рыб с хорошим зрением) видит не только предметы в окне (угол 97,6њ), но и предметы, отраженные водной поверхностью со дна (на рис. 2.11 это камни).
Механизм фоторецепции. Большой интерес представляет интимный механизм фоторецепции и трансформации энергии кванта света в энергию нервного импульса.
Чувствительность палочек и колбочек к свету обусловлена наличием в них пигментов. В химическом отношении любой из встречающихся в классе рыб зрительный пигмент является сложным белком, в составе которого имеется полипептидная структура, различная в палочках и колбочках, соединенная с одним из производных витамина А:
Родопсин = Ретиналь + палочковый белок опсин
Порфиропсин = Ретинен + палочковый белок опсин
Йодопсин = Ретиналь + колбочковый белок опсин
Цианопсин = Ретинен + колбочковый белок опсин
Молекулярную массу зрительных пигментов оценивают в 28 000-40 000, диаметр молекулы - в 40-50 ангстрем (А). В фоточувствительных клетках пигменты локализованы в мембранах наружных сегментов.
И ретиналь, и ретинен являются альдегидами витамина А. Они различаются только строением циклической части молекулы. У ретинена кольцо в положении "3" имеет дополнительную двойную связь (рис. 2.12).
Это различие приводит к смещению спектра поглощения пигмента в красную область. На действие кванта света реагирует 11-цисизомер ретиналя и ретинена. Под действием света углеродная цепь 11-цисизомера распрямляется и тянет за собой молекулу опсина, который изменяет свою конформацию

Рис. 2.11. Поле зрения рыбы
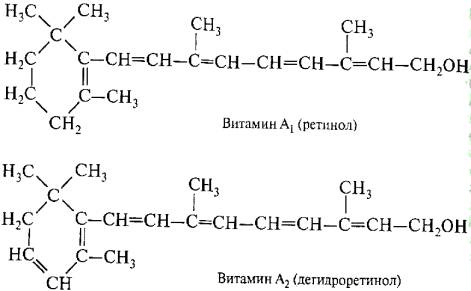
Рис. 2.12. Ретинол и ретинен (витамин А2)
Далее происходят два важных события.
Во-первых, светочувствительная клетка перемещается в пигментный слои сетчатки, где осуществляется восстановление изомера.
Во-вторых, конформационные изменения белка опсина приводят к изменению состояния мембраны эндоплазматического ретикулума с открыванием кальциевых каналов светочувствительных клеток.
В конечном счете изменяется мембранный потенциал базальной мембраны палочки (колбочки), которая является одновременно и пресинаптической частью синаптического образования с биполярной клеткой.
Палочки и колбочки имеют отрицательный потенциал покоя, реакция рецепции кванта света приводит к гиперполяризации клеточной мембраны, т. е. ее внутренняя сторона становится еще более электроотрицательной по отношению к наружной. Экспериментально доказано, что гиперполяризация мембраны не влияет на калиево-натриевый насос, однако изменяет мембранную проницаемость для ионов натрия.
Все эти электрохимические изменения фоторецепторов приводят к возбуждению биполярных клеток, а их деятельность в свою очередь, интегрируют ганглиозные клетки. Так рождается нервный импульс, затем поступающий в зрительные центры головного мозга.
Зрительный анализатор и развитие головного мозга. Уровень развития фоторецепции оказывает большое влияние на морфологию головного мозга, в частности развитие среднего мозга, мозжечка и ретикулярной формации ствола (рис. 2.13).
У активных дневных рыб - верховки и плотвы - хорошо развиты зрительные бугры среднего мозга, которые выполняют функцию конечной интеграции нервных импульсов, поступающих по зрительным нервам. У белуги, рыбы с сумеречным зрением, средний мозг развит хуже, зато хорошо развит обонятельный и продолговатый мозг, отвечающий за осязание. У рыб с острым зрением большой удельный вес в структуре головного мозга приходится на мозжечок. Возможно, это связано с тем, что рыбы с хорошим зрением, как правило, ведут активный образ жизни, т. е. обладают более сложными локомоторными реакциями.
У таких рыб, как щука, судак, окунь, лосось, на верхней проекции головного мозга средний мозг занимает 50-55 % площади проекции. У осетровых рыб площадь проекции среднего мозга составляет 13-23 %.
Роль зрительной сигнализации в онтогенезе существенно меняется. Параллельно меняется и морфология головного мозга. Например, мальки сазана длиной 7-10мм питаются планктоном, в поиске которого животные полагаются на зрение. Поэтому на этой стадии развития у сазана крупные глаза и хорошая острота зрения. Средний мозг на проекции головного мозга в это время занимает 45 % площади. У сазана, перешедшего на питание бентосом (длина 327мм), острота зрения падает, а проекция среднего мозга сокращается до 31 %. У взрослых рыб, ориентирующихся при поиске пище в основном на химическую и тактильную сигнализацию, этот показатель еще ниже.
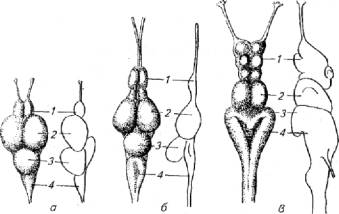
Рис. 2.13. Строение головного мозга трех видов рыб с различным зрением:
а -верховка, 6- плотва, в -белуга; 1- передний мозг; 2- средний мозг; 3, 4- задний мозг
Похоже, что и степень развития мозжечка у рыб связана с функцией зрения. Мозжечок хорошо развит у светолюбивых видов.
Таким образом, зрительный анализатор у рыб имеет большое значение. Зрение позволяет рыбам осуществлять адекватные реакции на изменения внешней среды. В процессе филогенеза развитие функции зрения стимулировало возникновение многих прогрессивных морфофункциональных адаптации и прежде всего развитие центральной нервной системы. В то же время следует подчеркнуть, что класс рыб достаточно разнообразен, и в нем имеется немало представителей, пищевая, половая, оборонительная и прочие виды активности которых не зависят или слабо зависят от зрения
§6. ХЕМОСЕНСОРИКА
Хеморецепция является древней формой взаимодействия организма со средой обитания. Вода служит одним из лучших растворителей химических веществ различной природы. Поэтов рыбы - обитатели водной среды - постоянно находятся в химических растворах. Естественно, что в процессе эволюции у рыб сформировался эффективный хеморецепторный аппарат, позволяющий им в своих целях использовать химическую информации из внешней среды.
Хеморецепция дает возможность рыбам отыскивать пищу, уходить от врагов, находить половых партнеров, узнавать представителей своего вида и своей стаи, наконец, отыскивать свои "исторические" места нереста и избегать неблагоприятных (по химическому составу воды) мест обитания.
Органы рыб, обеспечивающие восприятие и анализ химической информацией, условно подразделяют на органы обоняния, вкуса, обшей химической рецепции (Рис. 2.14).
Под обонянием рыб (ольфакторная рецепция) понимают рецепцию специфических веществ, находящихся в воде в малой концентрации. Обонятельные рецепторы обладают высокой чувствительностью и избирательностью и улавливают отдельные молекулы вещества в большом

Рис 2.14 Механизмы химической рецепции у рыб
объеме воды, т.е. являются дистантными рецепторами, позволяющими рыбам ориентироваться по запаху.
Контактные хеморецепторы возбуждаются при непосредственном соприкосновении с химическим веществом, например пищей. Этот тип рецепторов лежит в основе органов вкуса. Они возбуждаются сравнительно узким кругом химических веществ.
Кроме того, у рыб имеются малоспецифичные и низкочувствительные хеморецепторы, которые возбуждаются при раздражении веществами различной химической природы. Их часто называют рецепторами "общего химического чувства", призванными обеспечивать защитную реакцию на губительные для организма рыбы изменения химического состава среды обитания. Общее химическое чувство возникает при химическом раздражении свободных нервных окончаний тройничного, блуждающего и некоторых спинномозговых нервов.
Такое деление хеморецепторного аппарата рыб достаточно условно, поскольку, например, вкусовые ощущения складываются в результате афферентного синтеза импульсации, поступающей и от органов вкуса, и от органов обоняния. Пища с неприятным запахом невкусна, так же как и пища без ожидаемого запаха. Часто вкусовые ощущения рыб складываются в результате раздражения вкусовых почек и механорецепторов ротовой полости, головы, усиков и плавников.
В то же время неблагоприятные изменения химического состав водоеме (например, накопление аммиака, изменение раздражающие рецепторы общего химического чувства, искажают и вкусовые, и обонятельные ощущения у рыбы.
Обоняние рыб. Биологическое значение запахов в жизни рыб трудно переоценить. Химические раздражители воздействуют на рецепторы длительное время, т.е. не исчезают, как свет или звук. Пахучий след может сохраняться достаточно долго. Это важно при поиске пищи, обнаружении сородичей, а также уклонении от встречи с врагами.
У рыб хорошая память на запахи. Количество пахучих веществ (для рыб) велико, и они не поддаются классификации. В экспериментах рыбы успешно определяют возраст, стадию половой зрелости, иерархическое положение своего партнера, полагаясь исключительно на органы обоняния. Мигрирующие рыбы безошибочно определяют по химическому составу воду водоема, в котором они появились на свет.
Строение органов обоняния у рыб специфично и отражает водный образ жизни. К органам обоняния относят следующие парные (за исключением круглоротых) образования: ноздри, ольфакторные мешки, обонятельные нервы, обонятельные луковицы, у большинства видов рыб ноздри располагаются на верхней челюсти и ведут в ольфакторные (обонятельные) мешки.
Обонятельный мешок имеет сложное макро- и микростроение. Каждый мешок имеет вводное и выводное отверстия, через которые вода входит и после анализа удаляется из мешка. Дно мешка имеет складчатое строение, причем количество складок колеблется от двух до нескольких сотен. Кроме того, выделяют складки второго и третьего порядков. Есть мнение, что от количества складок (следовательно, обонятельной площади) зависит острота обоняния. Однако это обобщение справедливо для водных животных в целом. Среди рыб же имеются виды с хорошим обонянием, нос малым (всего 10-15) количеством складок (например, лососевые рыбы). У судака посредственное обоняние, но количество складок в 2-3 раза больше. Очевидно, что острота обоняния зависит и от микростроения слизистой оболочки ольфакторных мешков. Складки мешков имеют и еще одно назначение. Они обеспечивают определенную турбулентность воды при ее движении черв ольфакторные мешки. Интенсивность и характер циркуляции воды играют большую роль в процессе рецепции запахов.
Экспериментальные измерения показали, что угорь может менять скорость движения воды в мешках от 2 до 10 см/с.
Рыбы, с добавочными обонятельными мешками увеличивав скорость воды до 300 см/с даже при неподвижных челюстях.
Обонятельный эпителий, выстилающий ольфакторные мешки рыб, имеет сложную микроструктуру, отдаленно напоминающую микроструктуру сетчатки глаза (рис. 2.15).
Снаружи обонятельный эпителий покрыт двумя слоями слизи, под которыми находятся клетки эпителия. Основными рецепторными элементами принято считать биполярные нейроны трех типов: палочковидной, колбовидной и нитевидной форм. Однако не исследователи согласны с такой классификацией рецепторных клеток обонятельных клеток, считая, что в данном случае исследователи имеют дело с одним и тем же типом нейронов, находящихся в разном функциональном состоянии.

Рис. 2.15. Микроструктура эпителия:
1 и 3 - рецепторные клетки; 2 - большая слизистая клетка; 4 к 5- опорные клетки; 6- малая слизистая клетка
Не нашли, что искали? Воспользуйтесь поиском по сайту:
©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|

