|
|
Д) Взаимосвязь растительности с животным миромНе менее тесна взаимосвязь растительности с животным миром, населяющим данный биогеоценоз. Животные в процессе своей жизнедеятельности многосторонне влияют на растительность как непосредственно, питаясь ею, вытаптывая ее, строя в ней или при помощи ее свои жилища, убежища, содействуя опылению цветков и распространению семян или плодов, так и косвенно, изменяя почву, удобряя ее, разрыхляя, вообще меняя ее химические и физические свойства, а в некоторой степени влияя и на атмосферу. ПРИМЕР. Изучение отдельных комплексов вредителей определенных видов или других подсистем ценотического порядка (паразито-, микробоценозов и др.) в совокупности с растениями-"хозяевами", т.е. консорций, перспективно в прикладном отношении – для использования их в области защиты растений. Взаимоотношения между разными трофическими уровнями относятся к трофико-энергетическому направлению (Odum, 1963) и являются объектом многих исследований, получившим широкое развитие в последние десятилетия. Это дает возможность выявить общий характер и количественные показатели обмена веществ и энергии, раскрывая тем самым биогеофизическую и биогеохимическую роль живого покрова. Е) Взаимодействия между неживыми (абиотическими) компонентами Не только живые организмы взаимодействует с другими компонентами биогеоценоза, но и эти последние взаимодействуют друг с другом. Климатические условия (атмосфера) влияют на почвообразовательный процесс, а почвенные процессы, определяя выделение углекислоты и других газов (дыхание почвы), изменяют атмосферу. Почва влияет на животный мир, не только ее населяющий, но косвенно и на весь остальной животный мир. Животный же мир воздействует на почву. Факторы, влияющие на взаимодействия компонентов биогеоценоза Рельеф и биогеоценоз. Всякий биогеоценоз, занимая известное место в природе, связан с тем или иным рельефом. Но рельеф сам по себе не входит в число компонентов биогеоценоза. Рельеф является лишь условием, влияющим на процесс взаимодействия названных выше компонентов, и, в соответствии с этим, на их свойства и структуру, определяя направление и интенсивность процессов взаимодействия. В то же время взаимодействие компонентов биогеоценоза нередко может приводить к изменению рельефа и к созданию особых форм микрорельефа, а в известных случаях и мезо- и макрорельефа. Влияние человека на биогеоценоз.Человек не входит в число компонентов биогеоценозов. Он, однако, является в высшей степени мощным фактором, могущим не только в той или иной мере изменять, но и создавать путем культуры новые биогеоценозы. Ныне почти нет лесных биогеоценозов, не испытавших влияние хозяйственной, а часто и бесхозяйственной деятельности человека. Взаимовлияния между биогеоценозами. Вместе с тем каждый биогеоценоз, так или иначе, влияет на другие биогеоценозы и вообще явления природы, соседние с ним или в той или иной мере удаленные от него, т. е. обмен веществом и энергией имеет место не только между компонентами данного биогеоценоза, но и между самими фитоценозами. Часто ведущим фактором являются конкурентные отношения между фитоценозами. Фитоценоз более мощный вытесняет фитоценоз менее устойчивый, например, при определенных условиях происходит вытеснение соснового фитоценоза еловым, а вместе с этим изменяется и весь биогеоценоз. Таким образом, взаимодействие всех компонентов биогеоценоза, особенно лесного биогеоценоза (включая воду в почвогрунтах и атмосфере), очень разнообразно и сложно: - растительность все время находятся в зависимости от почвы, атмосферы, животного мира и микроорганизмов. - химический состав почвы, ее влага и физические свойства влияют на рост и развитие растений, на их плодоношение и возобновляемость, на технические свойства их древесины древесных пород, на их, на рост и развитие всей другой растительности. - вся растительность же в свою очередь в сильной степени воздействует на почву, определяя, главным образом, качество и количество органического вещества в почве, влияя на физические и химические ее особенности. Типы отношений между организмами в ценозе Организмы могут взаимодействовать друг с другом постоянно, в течение всей жизни или кратковременно. При этом они либо соприкасаются друг с другом, либо оказывают влияние на другой организм на расстоянии. Взаимные влияния растений могут иметь то благоприятныйдля их роста и развития характер, тонеблагоприятный. В первом случае условно говорят о «взаимопомощи», во втором – о «борьбе за существование» между растениями в широком, дарвинском смысле, или о конкуренции. Само собой разумеется, что все эти взаимовлияния между организмами в биоценозе в то же время играют большую роль и в биогеоценозе в целом. Они могут проходить между особями как разных видов, так и одного и того же вида, т. е. могут быть как межвидовыми, так и внутривидовыми. Отношения между организмами весьма многообразны. Удачна классификация эти отношений Г. Кларка (Clark, 1957) (табл. 1). Таблица 1 Классификация отношений между организмами(по Кларку, 1957)
Условные знаки: «+» – приращение или польза в жизненном процессе в результате отношений, «–» – уменьшение или ущерб, 0 – отсутствие заметного эффекта. Ниже рассматриваюся формы отношений, играющие наиболее существенную роль в фитоценозе. а) СИМБИОЗ ("ВЗАИМНЫЙ ПАРАЗИТИЗМ") – отношения между организмами, обычно разных видов и находящихся в более или менее длительном контакте, при которых один или оба организма извлекают из этих отношений пользу и ни один не испытывает ущерба. Первый тип симбиотических отношений, когда пользу извлекают оба организма, называется мутуализмом, второй, когда пользу извлекает лишь один из организмов, – комменсализмом («нахлебничеством»). Мутуализм Симбиоз азотфиксирующих организмов с голосеменными и цветковыми растениями – отношения между высшим растением и бактериями. На корнях многих растений встречаются клубеньки, образованные бактериями или реже грибами. Клубеньковые бактерии фиксируют атмосферный азот и переводят его в доступную для высших растений форму. ПРИМЕРЫ. Клубеньки на корнях растений из семейства бобовых образованы бактериями из рода ризобиум (Rhyzobium), а также на корнях видов лисохвоста, лоха, облепихи, подокарпуса, ольхи (Actinomyces alni) и других растений. Благодаря этому растения, зараженные клубеньковыми бактериями, могут хорошо расти на почвах, бедных азотом, а содержание азота в почве после культуры таких растений возрастает. В свою очередь бактерии получают от высших растений углеводы. Бактериальные желвачки с азотом были обнаружены не только на корнях, но и в листьях многих представителей семейств мирсиновых, диоскорейных, мареновых, примерно 370 видов растений. Бактерии, образующие эти клубеньки, проникают в листья через устьица еще тогда, когда листья свернуты в почках. Они фиксируют азот как клубеньковые бактерии, и отношения их с цветковыми растениями также носят характер взаимного паразитизма. Микориза – симбиотические отношения между высшим растением и грибом. Микоризы широко распространены среди диких и культурных растений. В настоящее время микориза известна более чем для 2000 видов высших растений (Федоров, 1954), но, несомненно, действительное число видов, для которых характерна микориза, значительно больше. Для высших растений, на корнях которых поселяются грибы, характерен особый тип питания – микотрофный. При микотрофном питании с помощью симбиотических грибов высшее растение получает зольные элементы пищи, в том числе азот, из органического вещества почвы. Что касается грибов, образующих микоризу, то они в большинстве не могут существовать без корневых систем высших растений, всасывающих влагу из почвы и поставляющих из кроны органическое вещество. Деревья растут гораздо лучше с микоризой, чем без нее. Различают два основных вида микоризы: эктотрофную и эндотрофную. При эктотрофной микоризе корень высшего растения окутывается плотным грибным чехлом, от которого отходят много численные гифы гриба. При эндотрофной микоризе мицелий гриба проникает в клетки корневой паренхимы корня, которые сохраняют свою жизнедеятельность. Промежуточную форму микоризы, при которой имеет место и наружное обрастание корня гифами гриба, и проникновение гифов внутрь корня, называют перитрофной (эктоэндотрофной), микоризой. Эктотрофная микориза – однолетняя. Она развивается летом или осенью и погибает к следующей весне. Она свойственна многим деревьям из семейств сосновых, буковых, березовых и др., а также некоторым травянистым растениям, например подъельнику. Эктотрофную микоризу чаще всего образуют базидиальные грибы из семейства Polyporaceae и особенно часто из рода Boletus. Так, подберезовик (В. scaber) образует микоризу на корнях березы, масленок – на корнях лиственницы (В. elegans) или сосны и ели (В. luteus), подосиновик (В. versipellis) – на корнях осины, белый гриб (В. edulus) – на корнях ели, дуба, березы (различные подвиды) и т. д. Эндотрофная микоризашироко распространена у растений семейств орхидных, вересковых, брусничных, а также у многолетних трав из семейства сложноцветных и у некоторых деревьев, на пример у красного клена (Acer rubrum) и др. В качестве второго компонента эндотрофной микоризы часто выступает гриб Phoma из группы несовершенных грибов. Эндотрофная микориза может быть образована Oreomyces (живет на корнях орхидей, по-видимому, может фиксировать азот) и некоторыми другими видами грибов. Как предполагалось ранее, этот гриб может усваивать азот из атмосферы. Это обстоятельство связано с тем, что вереск (Calluna) и другие представители семейства вересковых, а также виды семейства орхидных могут развиваться на безазотистой среде только в присутствии этого гриба. В отсутствие Phoma betake у этих растений не прорастают семена или гибнут проростки вскоре после прорастания семян. Гибель проростков у орхидей, грушанок и других лесных растений можно объяснить тем, что в их семенах почти полностью отсутствуют запасные питательные вещества в клетках, и поэтому без гифов гриба, доставляющих необходимые питательные вещества всходам, их развитие быстро прекращается. В сосновых лесах Центрального Предуралья (Логинова, Селиванов, 1968) насчитывается следующее содержание микотрофных видов в лесной микофлоре: в бору-беломошнике – 81%, в бору-брусничнике – 85, в бору-черничнике – 90, в бору сфагново-багульниковом – 45, в бору остепненном травяном – 89%. В пустынях Тау-Кум процент видов с микоризой в разных ассоциациях составляет от 42 до 69%. Значение микоризы в связи с ее широким распространением огромно. Многие растения – орхидные и вероятно вересковые, а также некоторые деревья без микоризы развиваются плохо или даже вообще не развиваются либо из-за недостатка питательных веществ в их мелких семенах, либо из-за недостаточного развития сосущих частей корешков, а также на бедных минеральными питательными веществами почвах. Грибы, образующие эндотрофную микоризу на их корнях, могут существовать лишь в кислой среде. Именно благодаря им многие представители орхидных и вересковых потому и живут только на кислых почвах. Следовательно, наличие в фитоценозе грибов, образующих микоризу, в значительной степени определяет видовой состав высших растений, входящих в этот фитоценоз, и служит немаловажным фактором в их борьбе за существование между растениями, поскольку отсутствие микоризы у растений, склонных к микотрофномy питанию, замедляет темпы их развития и ухудшает их положение по отношению к более быстро развивающимся видам, использующим микоризу. Коменсализм Наиболее характерные растения, которые можно привести в качестве примеров комменсализма по способу размещения в ценозе и по типу питания: эпифиты, лианы, почвенные и наземные сапрофиты.
Теневые эпифиты обитают в условиях сильной затененности, малого и мало меняющегося дефицита насыщения, т. е. в условиях, почти не отличающихся от условий жизни наземных трав. Они обитают главным образом в третьем (нижнем) ярусе леса. Многие из них имеют гигроморфную структуру тканей. Группа солнечных эпифитов, наиболее богатая по числу видов и особей, связана с кронами деревьев верхних ярусов. Эти эпифиты живут в микроклимате, промежуточном между микроклиматами напочвенного покрова и открытых мест, и получают гораздо больше света, чем теневые эпифиты. Многие солнечные эпифиты более или менее ксероморфны; осмотическое давление у них выше, чем у теневых эпифитов. Крайне ксерофильные эпифиты живут на верхушечных ветвях более высоких деревьев. Условия их местообитания подобны условиям открытых мест, условия питания здесь крайне суровы. Эпифиты, как правило, сапротрофы, т.е, питаются за счет отмирающих тканей растения-хозяина. Обычно для разложения этих отмирающих тканей эпифиты используют грибы, образующие микоризу с корнями эпифита. Большую роль в питании играют некоторые животные. ПРИМЕРЫ. Муравьи, поселяясь среди корней эпифитов, приносят в свои гнезда большое количество мертвых листьев, семян, плодов, которые, разлагаясь, дают питательные вещества эпифитам. Некоторые беспозвоночные и позвоночные животные поселяются в воде, скапливающейся в чашах, образованных листьями эпифитов из семейства бромелиевых, и их трупы, разлагаясь, дают пищу эпифитам. Наконец, среди эпифитов есть и насекомоядные растения, например виды рода непентес (Nepenthes) и некоторые пузырчатки.
В лесах умеренного пояса распространены исключительно или почти исключительно мелкие лианы, поэтому они не играют здесь большой роли. Почвенные и наземные сапрофиты. Сапрофиты – растительные организмы, живущие полностью (полные сапрофиты) или частично (частичные сапрофиты) за счет отмерших органов животных и растений. Кроме эпифитов, которые относятся по способу питания к сапрофитам, в эту группу входят многие наземные растения и обитатели почвы. ПРИМЕРЫ. К сапрофитам относится большинство грибов и бактерий, играющих огромную роль в круговороте веществ в почве, а также некоторые цветковые растения из семейств орхидных (гнездоцветка клобучковая) и вертляницевых (вертляница одноцветковая) в лесах умеренного пояса и из семейств лилейных, орхидных, горечавковых, истодовых и некоторых других в лесах тропического пояса. Большинство этих цветковых растений – полные сапрофиты, некоторые, по крайней мере, орхидеи, содержат какое-то количество хлорофилла и, вероятно, частично способны к фотосинтезу. Цвет надземных частей этих растений белый, светло-желтый, розовый, голубой или пурпурный. Обитают сапрофиты из цветковых растений в тропиках в тенистых местах на почве или на лежащих мертвых стволах. Обычно эти растения связаны с микоризными грибами, живущими на их корнях. Как правило, они невысоки, обычно не превышают 20 см, за исключением сапрофитной тропической орхидеи галеоны высочайшей (Gualala altissimo), представляющей собой лазающую (при помощи корней) лиану, достигающую высоты 40 м. Б) АНТАГОНИЗМ - отношения, при которых один или оба организма испытывают ущерб. Душители.Душители – корнесобственные растения, но начинают развитие в качестве эпифитов. Различные животные переносят их семена с одного дерева на другое. Птицы – основные переносчики семян душителей. Душитель образует корни двух родов: одни из них плотно прилегают к коре дерева-хозяина, ветвятся, и образуют плотную сеть, одевающую ствол дерева-хозяина, другие свешиваются вертикально вниз и, достигнув почвы, ветвятся в ней, доставляя душителю воду и минеральное питание. В результате затенения и сдавливания дерево-хозяин погибает, а душитель, к тому времени развивший мощный корневой «ствол», остается стоять на «собственных ногах». Многочисленные лианы фестонами свисают с дерева. Душители характерны для влажных тропиков. Душители находятся с деревьями-хозяевами в антагонистических отношениях. Некоторые южноамериканские виды-душители имеют настолько слабые корни, что при падении дерево-хозяин увлекает их за собой. Паразиты и полупаразиты.Паразитизм – отношения между организмами обычно разных видов, при которых один из них (паразит) проводит более или менее длительное время на теле или в теле другого организма (хозяина), получая питание из его тканей. Паразитические растения играют огромную роль в жизни фитоценозов, ослабляя одни растения, вызывая гибель других и тем самым, способствуют лучшему развитию третьих. Паразитов делят на две большие группы: факультативные и облигатные. Первые могут вести и паразитический, и сапрофитный образ жизни, как, например, грибы трутовики из рода полипорус (Polyporus), вторые ведут только паразитический образ жизни. В связи с этим облигатные паразиты обычно не сразу губят своего хозяина, а постепенно истощают его, в то время как факультативные паразиты весьма быстро приводят своего хозяина к гибели и переходят на питание его мертвыми тканями. Поэтому факультативные паразиты оказывают более существенное воздействие на растение-хозяина, чем облигатные. К числу паразитов относятся многие грибы, некоторые слизевики, водоросли и лишайники, а также ряд видов цветковых растений. Так, повилика европейская (Cuscuta europaea) иногда сильно поражает крапиву двудомную (Urtica dioica), а повилика хмелевидная (С. lupuliformis) и однопестичная (С. Monogyna) – иву (Salix). Паразитами следует считать и бактерии, вызывающие заболевания растений, животных и человека. Кроме полных паразитов, существуют гемипаразиты (полупаразиты). Последние, имея хлорофилл, способны к ассимиляции и от растения-хозяина получают преимущественно минеральное питание в водном растворе. Часть полупаразитов представляют виды, обычно обитающие на лугах, иногда в лесах, с очень слабыми корневыми системами. Эти растения присасываются своими корнями к корням цветковых растений. Это многие виды семейства норичниковых (Scrophulariaceae) из родов мытник (Pedicularis), марьянник (Melampyrum), зубянка (Odontites), очанка (Euphrasia), погремок (Alectorolophus) и др. Массовое их развитие может резко снизить качество луга, так как ценные в хозяйственном отношении травы сильно истощаются и резко снижают урожай. В условиях умеренного климата наиболее широко распространена омела белая (Viscum album) на лиственных, реже на хвойных породах. Хищничество – отношения между организмами разных видов (если организмы относятся к одному виду, то это каннибализм), при которых один из организмов (хищник) питается за счет второго организма (жертвы). Антибиоз – отношения между организмами, обычно относящимися к разным видам, при которых один из организмов причиняет вред другому (например, выделением вредных для другого организма веществ), не извлекая для себя из этих отношений видимого преимущества. Воздействие выделений одних растений на другие.Взаимоотношения между растениями, в которых ведущую роль играют специфически действующие продукты обмена веществ, Молиш (Molisch, 1937) назвал аллелопатией. Вещества, выделяемые надземными и подземными органами живых растений, и органические соединения, получающиеся в процессе разложения мертвых растительных остатков и оказывающие влияние на другие растения, называются колинами. Колины образуются как при свободной жизни растений рядом друг с другом, так и при паразитизме и взаимном влиянии пыльцы разных видов. Выделения веществ могут оказывать положительное или отрицательное влияние на растения того же вида или других видов. В частности, антибиотические вещества, препятствующие росту различных плесеней и бактерий, выделяют плесневые грибы, бактерии, актиномицеты, а также некоторые водоросли и лишайники. Эти выделения защищают растения от нападающих на них паразитов и тем самым ограничивают число видов, входящих в состав фитоценоза, или затрудняют развитие некоторых из них. Среди колинов различают: - газообразные выделения надземных органов растений, - прочие выделения наземных органов растений, - корневые выделения, - продукты распада мертвых растительных остатков. Среди газообразных выделений большую роль играет этилен, в значительных количествах вырабатываемый некоторыми растениями, например, яблоками. (Этилен задерживает рост, вызывает преждевременный опад листвы, ускоряет распускание почек и созревание плодов, оказывает положительное или отрицательное влияние на рост корней). Газообразные колины могут оказать воздействие на ход сезонных явлений в ценозе, а также подавить развитие тех или иных видов. Однако более или менее значительной роль газообразных колинов может быть лишь в аридных регионах, где обильно число растений, выделяющих различные легко испаряющиеся эфирные масла. Эти эфирные масла служат приспособлением к понижению температуры вокруг испаряющей поверхности, но одновременно могут оказывать определенное воздействие на те или иные растения. Твердые и жидкие выделения надземных органов растений представляют собой минеральные и сложные органические соединения, которые вымываются из надземных частей растений осадками, иногда в весьма значительных количествах, и оказывают действие на другие растения, попадая на них непосредственно с дождем, росой или через почву, куда они вмываются. ПРИМЕРЫ. Выделения полыни горькой (Artemisia absinthium) задерживают рост многих растений, то же указывается для веществ, содержащихся в листьях ореха черного (Juglans nigra), а также в листьях и хвое многих древесных пород и некоторых кустарников и трав. Ингибирующим влиянием у дальневосточных видов обладает злак вейник Лангсдорфа, возможно, существуют какие-то выделения у волжанки двудомной и винограда амурского. В то же время известно благотворное влияние на прорастание семян хвойных вытяжки из брусники, зеленых мхов. Конкуренция – вслед за Ч. Дарвином в широком смысле – это борьба за существование: борьба за пищу, за место или за какие-либо другие условия. Даже при достаточно высоком сходстве экологических требований, растения одних видов оказываются более сильными, более конкурентомощными при одних определенных значениях факторов среды, другие – при других. Это и служит причиной победы того или другого вида в межвидовой борьбе. ПРИМЕР. На Крайнем Севере Дальнего Востока береза каменная, ольховый и кедровый стланики формируют чистые сообщества и сообщества с доминированием одного из них на склонах южных экспозиций. Часто они произрастают совместно и доминанта трудно выделить. Все три вида характеризуются очень близкими экологическими свойствами. Все они – реликтовые, и отличаются высоким тепло-, влаго- и светолюбием. Но при этом ольховник несколько более теневынослив и более требователен к влажности почв, береза – более требовательна к теплу и трофности почв, а кедровый стланик – к свету и влажности воздуха. В результате при совместном произрастании кедровостланиковые ценоэлементы, или парцеллы, обычно приурочены к повышенным элементам микрорельефа, более сухим и хорошо дренированным, каменноберезовые – к выровненным участкам с высокой трофностью и тоже хорошо дренированных, ольховниковые – к микропонижениям с очень высоким, но проточным увлажнением и высокой трофностью почв. Каменноберезняки чаще приурочены к распадкам и в горах не поднимаются выше стлаников, кедровый стланик образует чистые заросли на верхней границе леса и на грядах, расположенных полосами вдоль по склону, а заросли ольховника предпочитают седловины и перегибы поверхностей склона в местах с вогнутой поверхностью. Конкуренция отмечается между особями одного вида (внутривидовая борьба) и между особями разных видов (межвидовая борьба) в неблагоприятных условиях среды. Оба типа борьбы обычно тесно связаны друг с другом. В борьбе за пищу, за влагу, с вредителями и паразитами растения конкурируют и с особями своего вида, и с особями других видов.
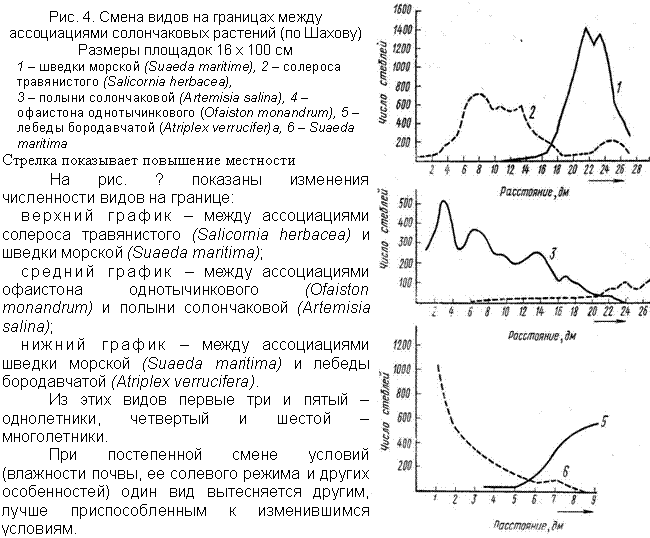
В каждом фитоценозе подбираются растения: - представляющие различные жизненные формы и занимающие место в различных синузиях, ярусах, микроценозах, т.е. образующие группы, характеризующиеся неодинаковым отношением к среде и неодинаковым местом в фитоценозе; - дифференцированные по срокам прохождения сезонных фаз. Сочетание в одном фитоценозе растений с различными экологическими особенностями – тенелюбивых и светолюбивых, в различной степени приспособленных к недостатку влаги и другим факторам среды, позволяет фитоценозу наиболее полно использовать условия местообитания. Смена видов происходит не сразу, постепенно один вид вытесняет другой, поэтому между фитоценозами обычно нет четкой границы. Полоса на которой происходит смена фитоценозов, называется экотоном. В экотоне, как правило, присутствуют виды смежных сообществ, и мозаичность растительного покрова здесь выше, но жизненное состояние видов-доминантов обоих сообществ в экотоне обычно хуже, чем в тех ценозах, условия которых этим видам соответствуют больше. Вытеснение одних видов другими на границе фитоценозов (правда, не одновидовых) происходит и без изменений условий среды, в результате различной конкурентной способности видов, в частности различной энергии вегетативного размножения. ПРИМЕРЫ. Так, известный всем сорный злак пырей не только способен заглушить культурные посевы, но и вытесняет многие дикие виды (крапиву, чистотел и др.), растущие с ним по соседству и которые вегетативно размножаются очень слабо. Даже клевер ползучий постепенно уступает позиции пырею. Очень сильной конкурентной способностью обладает мох сфагнум. По мере разрастания он в прямом смысле поглощает соседние растения. В районах распространения вечной мерзлоты фитоценозы с господством сфагнума занимают обширные пространства, вытесняя зоны своего влияния не только травы и кустарнички, но и кустарники и деревья. В результате борьбы за существование происходит дифференциация видов, образующих фитоценоз. В то же время строение фитоценоза – не только результат борьбы за существование, но и результат приспособления растений к уменьшению напряженности этой борьбы. В фитоценозе виды подобраны таким образом, что дополняют друг друга своими свойствами. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|