|
|
Железы внутренней секреции
Железы внутренней секреции, как и гормоны, выделяемые ими, имеют разное происхождение, что важно для изучения их эволюции. Некоторые эндокринные железы связаны по происхождению с эпителиальной выстилкой глотки. К ним относятся щитовидная и паращитовидная железы. Эпифиз развивается как вырост мозга; гипофиз, надпочечники и поджелудочная железа имеют сложное происхождение. Среди хордовых только у бесчерепных эндокринная система существует в виде отдельных клеток и клеточных комплексов, которые находятся в разных отделах тела, объединенных друг с другом за счет гуморального взаимодействия. У позвоночных в основании промежуточного мозга развивается гипоталамус — нейросекреторное образование, осуществляющее связь между двумя системами интеграции организма в единое целое: нервной и эндокринной. Вместе с гипофизом гипоталамус образует единую гипоталамо-гипофизарную систему. Эволюционным предшественником гипоталамуса является так называемый инфундибулярный вырост ланцетника, состоящий из нейросекреторных клеток и находящийся на вентральной стороне переднего конца нервной трубки. Начиная с рыб гипоталамус дифференцируется на многочисленные ядра, клетки которых с помощью отростков контактируют как с нейронами мозга, так и с клетками гипофиза. Нейросекреторные клетки гипоталамуса выделяют две основные группы гормонов: пептидные и моноаминовые. Первые — гормоны, влияющие на функции внутренних органов — вазопрессин, регулирующий артериальное давление, окситоцин, действующий на мускулатуру матки, и др. Вторая группа гормонов (дофамин, норадреналин, серотонин) регулирует деятельность передней доли гипофиза. Под их действием стимулируется или подавляется секреция гормонов соответствующими гипофизарными клетками. Подробнее остановимся на эволюции гипофиза. Эта железа состоит из трех долей: передней (аденогипофиза), средней (промежуточной) и задней (нейрогипофиза). Гипофиз соединен с гипоталамусом его выростом — воронкой, через которую проходят отростки нейронов гипоталамуса и кровеносные сосуды, обеспечивающие их гуморальное взаимодействие. Доли гипофиза имеют разное происхождение. Передняя доля развивается из выпячивания эктодермального эпителия крыши ротовой полости, так называемого кармана Ратке, который растет в сторону промежуточного мозга. Задняя доля развивается из задней части воронки. Клетки, входящие в ее состав, по происхождению являются глиальными. Промежуточная доля — производная от передней (рис. 14.42).
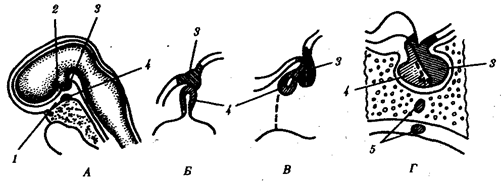
Рис. 14.42. Развитие гипофиза у человека. А—сагиттальный срез 4-недельного зародыша; Б—объединение зачатков гипофиза из стенки глотки и основания мозга; В—формирование трех долей гипофиза на 8-й неделе развития; Г—сформированный гипофиз:
1—крыша ротовой полости, 2—основание мозга, 3—выпячивание основания мозга (задняя доля гипофиза), 4—карман Ратке (промежуточная и передняя доли гипофиза), 5—аномалия развития (добавочные дольки передней доли в стенке глотки и внутри клиновидной кости, оставшиеся в области выпячивания кармана Ратке)
У хрящевых рыб во взрослом состоянии сохраняется первоначальная связь передней доли гипофиза с эпителием ротовой полости. За счет ее задней части формируется также средняя доля. Обе доли вырабатывают гонадотропные гормоны. У костных рыб и личинок земноводных имеются передняя и промежуточная доли, а у взрослых амфибий, переходящих к наземному существованию, появляется также задняя, регулирующая водный обмен. Средняя доля у них перестает выделять гонадотропный гормон, но вырабатывает пролактин. В связи с наземным образом жизни пресмыкающихся и млекопитающих у них наиболее прогрессивно развивается задняя доля гипофиза, что связано с интенсификацией водного обмена. Передняя доля вырабатывает соматотропный гормон (гормон роста) и ряд гормонов, регулирующих функции других желез внутренней секреции, а средняя — пролактин и некоторые другие. При этом усиливается дифференцировка и интенсифицируются функции ядер гипоталамуса, находящихся в функциональной связи со всеми долями гипофиза. У человека в эмбриогенезе развитие гипофиза соответствует основным этапам его эволюции. Очень часто, в 30—40%, у нормальных людей под слизистой оболочкой крыши глотки, в основании клиновидной кости, обнаруживается группа клеток длиной 5—6 мм и шириной 0,5—1 мм, по структуре и функциям соответствующая передней доле гипофиза. Это результат нарушения перемещения клеток при закладке гипофиза в эктодерме ротовой полости в области турецкого седла. Эту 139 аномалию называют эктопией аденогипофиза, она не сопровождается патологическими проявлениями. Более опасно сохранение полости в области кармана Ратке — киста кармана Ратке. Она располагается между передней и промежуточной долями, содержит слизь и в ряде случаев имеет тенденцию к росту и даже к переходу в злокачественное новообразование. Растущие кисты сдавливают гипофиз и вызывают его гипофункцию. Больные нуждаются в оперативном вмешательстве. Щитовидная железа, гормон которой — тироксин — регулирует энергетический обмен, среди хордовых как компактный орган впервые появляется у рыб. Однако уже у ланцетника отдельные тироксин-синтезирующие клетки обнаруживаются в желобке на вентральной стороне глотки. Щитовидная железа рыб закладывается также в виде желобка на вентральной стороне глотки между 1-й и 2-й жаберными щелями в области зачатка основания языка. Позже этот клеточный материал погружается под слизистую оболочку и формирует фолликулы, характерные для щитовидной железы. У других позвоночных железа закладывается так же, как у рыб, но затем она перемещается в область подъязычной кости (у земноводных) или в шейную область (у пресмыкающихся и млекопитающих). У человека в эмбриогенезе щитовидной железы происходит рекапитуляция предковых состояний. Гетеротопия ее осуществляется посредством миграции клеток в виде тяжа, полого внутри, называющегося щитоязычным протоком — canalis thyreoglossus. При нормальном развитии этот проток полностью редуцируется, рудиментом его является слепое отверстие foramen caecum в корне языка. Свидетельством эмбрионального перемещения железы является и расположение верхней щитовидной артерии a. thyreoidea superior, которая, начинаясь от общей сонной артерии, резко поворачивает вниз и входит в ее ткань. На ранних этапах эмбрионального развития эта артерия направляется вверх к зачатку щитовидной железы, а затем меняет свое направление вместе с его перемещением книзу. Персистирование участков подъязычного протока в постнатальном периоде сопровождается накоплением в них жидкости и образованием срединных кист шеи, которые могут располагаться в любом месте от корня языка до верхней границы щитовидного хряща. Иногда, кисты загнаиваются и прорываются с образованием срединных свищей шеи. Не менее известны такие пороки развития, как эктопия щитовидной железы, которая может развиться в результате нарушения клеточной миграции (рис. 14.43). Из эпителия глотки в области III-V жаберных карманов у позвоночных развиваются мелкие эндокринные образования, связанные по месту окончательного положения с щитовидной железой. Это паращитовидные железы и ультимобранхиальные тельца. Первые, выделяя гормон паратиреоидин, повышающий содержание ионов кальция в крови и уменьшающий их количество в костях, развиваются как самостоятельные железы только у наземных позвоночных, а у земноводных — лишь после метаморфоза. Клетки вторых выделяют кальцитонин, являющийся антагонистом паратиреоидина. Таким образом, паращитовидные железы и ультимобранхиальные тельца являются регуляторами кальциевого обмена. Значение их наиболее велико у наземных позвоночных. В филогенетическом ряду позвоночных они постепенно перемещаются из глоточной области в сторону щитовидной железы, а у млекопитающих даже срастаются с ней. Паращитовидные железы еще сохраняют самостоятельность, а клетки ультимобранхиальных телец мигрируют между фолликулами щитовидной железы и сохраняются под названием парафолликулярных клеток. Таким образом, на примере щитовидной, паращитовидных желез и ультимобранхиальных телец видна интеграция железистых структур в сложное надерганное образование, выполняющее целый комплекс взаимосвязанных функций.
Рис. 14.43. Аномалии щитовидной железы: 1—нормальное расположение. железы, 2—место эмбриональной закладки железы, 3—варианты аномального расположения железы; стрелками показано направление перемещения зачатка щитовидной железы в эмбриогенезе
У человека известны аномалии расположения паращитовидных желез, связанные с нарушением их гетеротопий. Надпочечники позвоночных имеют двойственное происхождение. У рыб и земноводных ткани, соответствующие мозговому и корковому веществам этих желез, расположены отдельно друг от друга. Зачатки мозгового вещества связаны по происхождению с симпатическими нервными узлами и расположены метамерно. Зачатки коркового вещества развиваются из утолщений эпителия брюшины. У наземных позвоночных мозговое и корковое вещества объединяются в компактные эндокринные железы, имеющие сложное гистологическое строение. У млекопитающих они прилежат к переднему концу почек. Мозговое вещество выделяет в основном адреналин — регулятор кровообращения и энергетического обмена, а корковое вещество — разнообразные стероидные гормоны, влияющие на минеральный, углеводный обмены и функции почек. Эндокринная система. В любом организме вырабатываются соединения, разносящиеся по всему организму, имеющие интегративную роль. У растений есть фитогормоны, контролирующие рост, развитие плодов, цветов, развитие пазушных почек, деление камбия и др. Фитогормоны есть у одноклеточных водорослей. Гормоны появились у многоклеточных организмов, когда возникли специальные эндокринные клетки. Однако химические соединения, играющие роль гормонов, были и раньше. Тироксин, трийодтиронин (щитовидная железа) обнаружены у цианобактерий. Гормональная регуляция у насекомых изучена плохо. В 1965 году Вильсон выделил инсулин из морской звезды. Оказалось, что дать определение гормону очень трудно. Гормон – это специфическое химическое вещество, выделяемое особыми клетками в определенном участке тела, которое поступает в кровь и затем оказывает специфическое действие на определенные клетки или органы-мишени, расположенные в других областях тела, что приводит к координации функций всего организма в целом. Известно большое количество гормонов млекопитающих. Они делятся на 3 основные группы. Феромоны. Выделяются во внешнюю среду. С их помощью животные принимают и передают информацию. У человека запах 14 - окситететрадекановой кислоты четко различают только женщины, достигшие половой зрелости. Наиболее просто организованные многоклеточные организмы – например, губки тоже имеют подобие эндокринной системы. Губки состоят из 2 слоев – энтодермой и экзодермой, между ними располагается мезенхима, в которой содержатся макромолекулярные соединения, характерные для соединительной ткани более высокоорганизованных организмов. В мезенхиме есть мигрирующие клетки, некоторые клетки способны секретировать серотонии, ацетилхолин. Нервная система у губок отсутствует. Вещества, синтезируемые в мезенхиме, служат для связи отдельных частей организма. Координация осуществляется за счет перемещения клеток по мезенхиме. Есть также и перенос веществ между клетками. Заложена основа химической сигнализации, которая характерна для остальных животных. Самостоятельных эндокринных клеток нет. У кишечнополостных имеется примитивная нервная система. Первоначально нервные клетки выполняли нейросекреторную функцию. Трофическую функцию, осуществляли контроль роста, развития организма. Затем нервные клетки стали вытягиваться и образовали длинные отростки. Секрет выделялся около органа-мишени, без переноса (т.к. не было крови). Эндокринный механизм возник раньше проводникового. Нервные клетки были эндокринными, а потом получили и проводниковые свойства. Нейросекреторные клетки был первыми секреторными клетками. Первичноротые и вторичноротые вырабатывают одинаковые стероидные и пептидные гормоны. Принято считать, что в процессе эволюции из одних полипептидных гормонов могут возникнуть новые (мутации, дупликации генов). Дупликации менее подавляются естественным отбором, чем мутации. Многие гормоны могу синтезироваться не в одной железе, а в нескольких. Например, инсулин вырабатывается в поджелудочной железе, подчелюстной железе, 12-перстной кишке и других органах. Существует зависимость генов, контролирующих синтез гормонов от положения. 103.Филогенез иммунной систем.Эволюция живого мира состояла в образовании таких форм жизни, которые активно взаимодействуют с другими живыми организмами. Биологические организмы существуют в биотическом окружении. Они взаимодействуют с другими организмами, в биоценозе существует круговорот. Абиотические факторы менее сильно воздействуют на организм. Эволюция живой материи привела к возникновению динамической системы взаимозависимых организмов, не могут существовать без биотического взаимодействия. Такие взаимодействия проявляются в питании и обмене веществ. Существует тенденция к усложнению связей. Современные многоклеточные организмы во многом утратили возможность синтезировать даже простые органические вещества, но научились добывать их в готовом виде. Это консументы (в том числе человек). Жизнь основана на питании продуктами биосинтеза, создание которых осуществляется продуцентами (зеленые растения). Жизнь консументов напрямую зависит от биологической продуктивности растений и от развитости растениеводства. Растениеводство- источник пищи и сырья для промышленности (в том числе фармацевтической промышленности). 40% лекарственных средств в мировой фармацевтической промышленности растительного происхождения, снижение продуктивности растений приводит к голоданию, недоеданию, ухудшению качества жизни, снижению производства и уровня общественного здоровья. Важнейший фактор, понижающий урожайность растений – микроорганизмы, грибы, бактерии. Они приспособлены к паразитизму на разных растениях, животных и человеке. Пандемия гриппа – «испанки» унесла жизни 20 млн. человек. Жертвы микробного паразитизма - все живые организмы, в том числе и сами микробы. Почему земле до сих пор не населена одними микроорганизмами? Почему жестокий антагонизм между живыми существами не привел к гибели живой материи еще на заре эволюции? Почему эволюция не остановилась? Ответы на эти и многие другие вопросы дает иммунология. Иммунитет – невосприимчивость, устойчивость, резистентность, толерантность – способность организма противостоять агрессии со стороны других биологических видов. 1902 г. Мечников и Зильбер основали учение об иммунитете. «Иммунитет – это совокупность всех наследственных полученных и индивидуально приобретенных организмом свойств, которые препятствуют проникновению и размножению микроорганизмов, вирусов и других патогенных объектов и действию выделяемых ими продуктов». Наследственный иммунитет – свойства невосприимчивости, полученные от предков. Он определяется генотипом. Индивидуальный иммунитет - приобретенный иммунитет, который вырабатывается на протяжении жизни организма. Выделяют следующие группы факторов иммунитета: - фагоцитарные; - конституциональные; - лимфоидные. Лимфоидные факторы – наследственная способность организмов создавать иммунитет. Конституциональные – имеют общебиологическое значение. Они присутствуют у всех организмов независимо от таксономического положения. У беспозвоночных и позвоночных существует система фагоцитоза. У растений и простейших подобная система отсутствует. Растения защищены только наследственными свойствами. Грибы и вирусы тоже не защищены. Только у позвоночных существует приобретенная невосприимчивость благодаря наличию лимфоидной системы. При этом защитные функции лимфатической системы осуществляются специфической активностью антител и иммуноглобулинов. Антигены – любые вещества, удовлетворяющие требованиям: - индицируют образование соответствующих антител при введении в организм; - вступают в реакцию с молекулой комплементарного иммуноглобулина. Наибольшей антигенной активностью обладают белки, меньшей – полисахариды и нуклеиновые кислоты. Существует большое количество антител. Антитела вырабатываются комплексом органов: - вилочковая железа; - селезенка; - кровь; - пейеровы бляшки тонкого кишечника; - костный мозг; - сумка Фабрициуса (у птиц). Изучение иммунитета производится на растениях. Восприимчивость генетически детерминирована (доминантные и рецессивные признаки). Более 50 лет изучается иммунитет иммуногенетикой. 104.Сравнительный обзор скелета позвоночных животных. Скелет головы. Осевой скелет. Скелет конечностей. Основные тенденции прогрессивной эволюции. Врожденные пороки развития скелета у человека. Филогенез двигательной функции лежит в основе прогрессивной эволюции животных. Поэтому уровень их организации в первую очередь зависит от характера двигательной активности, которая определяется особенностями организации опорно-двигательного аппарата, претерпевшего в типе Хордовые большие эволюционные преобразования в связи со сменой сред обитания и изменения форм локомоции. Действительно, водная среда у животных, не имеющих наружного скелета, предполагает однообразные движения за счет изгибов всего тела, в то время как жизнь на суше более способствует их перемещению с помощью конечностей. Рассмотрим в отдельности эволюцию скелета и мышечной системы. Скелет
У хордовых скелет внутренний. По строению и функциям подразделяется на осевой, скелет конечностей и головы. Осевой скелет
В подтипе Бесчерепные имеется только осевой скелет в виде хорды. Она построена из сильно вакуолизированных клеток, плотно прилегающих друг к другу и покрытых снаружи общими эластической и волокнистой оболочками. Упругость хорде придают тургорное давление ее клеток и прочность оболочек. Хорда закладывается в онтогенезе всех хордовых и выполняет у более высокоорганизованных животных не столько опорную, сколько морфогенетическую функцию, являясь органом, осуществляющим эмбриональную индукцию. На протяжении всей жизни у позвоночных хорда сохраняется только у круглоротых и некоторых низших рыб. У всех остальных животных она редуцируется. У человека в постэмбриональном периоде сохраняются рудименты хорды в виде nucleus pulposus межпозвоночных дисков. Сохранение избыточного количества хордального материала при нарушении его редукции чревато возможностью развития у человека опухолей — хордом, возникающих на его основе. У всех позвоночных хорда постепенно вытесняется позвонками, развивающимися из склеротомов сомитов, и функционально заменяется позвоночным столбом. Это один из выраженных примеров гомотопной субституции органов (см. § 13.4). Формирование позвонков в филогенезе начинается с развития их дуг, охватывающих нервную трубку и становящихся местами прикрепления мышц. Начиная с хрящевых рыб обнаруживается охрящевение оболочки хорды и разрастание оснований позвонковых дуг, в результате чего формируются тела позвонков. Срастание верхних позвонковых дуг над нервной трубкой образует остистые отростки и позвоночный канал, в который заключена нервная трубка (рис. 14.6).
Рис. 14.6. Развитие позвонка. А—ранний этап; Б—последующая стадия: 1—хорда, 2—оболочка хорды, 3—верхние и нижние позвонковые дуги, 4—остистый отросток, 5—зоны окостенения, 6—рудимент хорды, 7—хрящевое тело позвонка
Замещение хорды позвоночным столбом — более мощным органом опоры, имеющим сегментарное строение,— позволяет увеличить общие размеры тела и активизирует двигательную функцию. Дальнейшие прогрессивные изменения позвоночного столба связаны с тканевой субституцией — заменой хрящевой ткани на костную, что обнаруживается у костных рыб, а также с дифференцировкой его на отделы. У рыб только два отдела позвоночника: туловищный и хвостовой. Это связано с перемещением их в воде за счет изгибов тела. Земноводные приобретают также шейный и крестцовый отделы, представленные каждый одним позвонком. Первый обеспечивает большую подвижность головы, а второй — опору задним конечностям. У пресмыкающихся удлиняется шейный отдел позвоночника, первые два позвонка которого подвижно соединены с черепом и обеспечивают большую подвижность головы. Появляется поясничный отдел, еще слабо отграниченный от грудного, а крестец состоит уже из двух позвонков. Млекопитающие характеризуются стабильным количеством позвонков в шейном отделе, равным 7. В связи с большим значением в движении задних конечностей крестец образован 5—10 позвонками. Поясничный и грудной отделы четко отграничены друг от друга. У рыб все туловищные позвонки несут ребра, не срастающиеся друг с другом и с грудиной. Они придают телу устойчивую форму и обеспечивают опору мышцам, изгибающим тело в горизонтальной плоскости. Эта функция ребер сохраняется у всех позвоночных, совершающих змеевидные движения,— у хвостатых земноводных и пресмыкающихся , поэтому у них ребра также располагаются на всех позвонках, кроме хвостовых. У пресмыкающихся часть ребер грудного отдела срастается с грудиной, формируя грудную клетку, а у млекопитающих в состав грудной клетки входит 12—13 пар ребер.
Рис. 14.7. Аномалии развития осевого скелета. А — рудиментарные шейные ребра (показаны стрелками); Б — несращение остистых отростков позвонков в грудной и поясничной областях. Спинномозговые грыжи
Онтогенез осевого скелета человека рекапитулирует основные филогенетические стадии его становления: в периоде нейруляции закладывается хорда, заменяющаяся впоследствии хрящевым, а затем и костным позвоночником. На шейных, грудных и поясничных позвонках развивается по паре ребер, после чего шейные и поясничные ребра редуцируются, а грудные срастаются спереди друг с другом и с грудиной, формируя грудную клетку. Нарушение онтогенеза осевого скелета у человека может выразиться в таких атавистических пороках развития, как несрастание остистых отростков позвонков, в результате чего формируется spinabifida — дефект позвоночного канала. При этом часто через дефект выпячиваются мозговые оболочки и образуется спинномозговая грыжа (рис. 14.7). В возрасте 1,5—3 мес. зародыш человека обладает хвостовым отделом позвоночника, состоящим из 8—11 позвонков. Нарушение их редукции в последующем объясняет возможность возникновения такой известной аномалии осевого скелета, как персистирование хвоста. Нарушение редукции шейных и поясничных ребер лежит в основе их сохранения в постнатальном онтогенезе.
Скелет головы
Продолжением осевого скелета спереди является осевой, или мозговой, череп, служащий для защиты головного мозга и органов чувств. Рядом с ним развивается висцеральный, или лицевой череп, образующий опору передней части пищеварительной трубки. Обе части черепа развиваются по-разному и из разных зачатков. На ранних этапах эволюции и онтогенеза они не связаны между собой, но позже эта связь возникает.
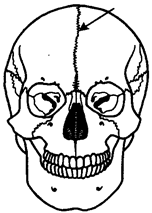
Рис. 14.8. Череп человека с методическим швом (указан стрелкой)
В задней части осевого черепа в процессе развития обнаруживаются следы сегментации, поэтому считают, что он представляет собой результат слияния друг с другом закладок передних позвонков. В состав мозгового черепа включаются также закладки хрящевых капсул мезенхимального происхождения, окружающие органы слуха, обоняния и зрения. Кроме того, часть мозгового черепа (лежащая кпереди от турецкого седла), не имеющая сегментации, развивается, по-видимому, как новообразование в связи с увеличением размеров переднего мозга. Филогенетически мозговой череп прошел три стадии развития: перепончатую, хрящевую и костную. У круглоротых он практически весь перепончатый и не имеет передней, несегментированной, части. Череп хрящевых рыб почти полностью хрящевой, причем включает в себя как заднюю, первично сегментированную, часть, так и переднюю. У костных рыб и остальных позвоночных осевой череп становится костным за счет процессов окостенения хряща в области его основания (основная, клиновидная, решетчатая кости) и за счет возникновения покровных костей в верхней его части (теменные, лобные, носовые кости). Кости осевого черепа в процессе прогрессивной эволюции претерпевают олигомеризацию. Появление большого количества зон окостенения и последующее слияние их вместе при формировании таких костей, как лобная, височная и др., свидетельствуют об этом. Широко известны у человека такие аномалии мозгового черепа, как наличие межтеменных, а также двух лобных костей с метопическим швом между ними (рис. 14.8). Никакими патологическими явлениями они не сопровождаются и обнаруживаются поэтому обычно случайно после смерти. Висцеральный череп впервые появляется также у низших позвоночных. Он формируется из мезенхимы эктодермального происхождения, которая группируется в виде сгущений, имеющих форму дужек, в промежутках между жаберными щелями глотки. Первые две дужки получают особенно сильное развитие и дают начало челюстной и подъязычной дугам взрослых животных. Следующие дуги в числе 4—5 пар выполняют опорную функцию для жабр и называются жаберными. У хрящевых рыб впереди челюстной дуги располагаются обычно еще 1—2 пары предчелюстных дуг, имеющих рудиментарный характер. Это свидетельствует о том, что у предков позвоночных имелось большее количество висцеральных дуг, чем б или 7, а их дифференцировка происходила на фоне олигомеризации. Челюстная дуга состоит из двух хрящей. Верхний называют нёбно-квадратным, он выполняет функцию первичной верхней челюсти. Нижний, или меккелев, хрящ — первичная нижняя челюсть. На вентральной стороне глотки меккелевы хрящи соединены друг с другом таким образом, что челюстная дуга кольцом охватывает ротовую полость. Вторая висцеральная дуга с каждой стороны состоит из гиомандибулярного хряща, сращенного с основанием мозгового черепа, и гиоида, соединенного с меккелевым хрящом. Таким образом, у хрящевых рыб обе первичные челюсти соединены с осевым черепом через вторую висцеральную дугу, в которой гиомандибулярный хрящ выполняет роль подвеска к мозговому черепу. Такой тип соединения челюстей и осевого черепа называют гиостильным (рис. 14.9). У костных рыб начинается замещение первичных челюстей вторичными, состоящими из накладных костей — челюстной и предчелюстной сверху и зубной внизу. Нёбно-квадратный и меккелев хрящи при этом уменьшаются в размерах и смещаются кзади. Гиомандибулярный хрящ продолжает выполнять функции подвеска, поэтому череп остается гиостильным. Земноводные в связи с переходом к наземному существованию претерпели значительные изменения висцерального черепа. Жаберные дуги частично редуцируются, а частично, меняя функции, входят в состав хрящевого аппарата гортани. Челюстная дуга своим верхним элементом — нёбно-квадратным хрящом — срастается полностью с основанием мозгового черепа, и череп становится, таким образом, аутостильным. Гиомандибулярный хрящ, сильно редуцированный и освободившийся от функции подвеска, располагаясь в области первой жаберной щели внутри слуховой капсулы, взял на себя функцию слуховой косточки — столбика,— передающей звуковые колебания от наружного к внутреннему уху. Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Часть хрящевого материала жаберных дуг входит в состав не только гортани, но и трахеи. Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения. Одна слуховая косточка — столбик,— характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко, а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку и молоточек. Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих (рис. 14.9).
Рис. 14.9. Эволюция двух первых висцеральных жаберных дуг позвоночных. А—хрящевая рыба; Б—земноводное; В—пресмыкающееся; Г— млекопитающее: 1—нёбно-квадратный хрящ, 2—меккелев хрящ, 3—гиомандибулярный хрящ, 4—гиоид, 5—столбик, 6—накладные кости вторичных челюстей, 7—наковаленка, 8—стремечко, 9—молоточек; гомологичные образования обозначены соответствующей штриховкой Рекапитуляция основных этапов филогенеза висцерального черепа происходит и в онтогенезе человека. Нарушение дифференцировки элементов челюстной жаберной дуги в слуховые косточки является механизмом формирования такого порока развития среднего уха, как расположение в барабанной полости только одной слуховой косточки — столбика, что соответствует строению звукопередающего аппарата земноводных и пресмыкающихся. Скелет конечностей
У хордовых выделяются непарные и парные конечности. Непарные (спинные, хвостовой' и анальный плавники) являются основными органами передвижения у бесчерепных, рыб и в меньшей степени у хвостатых амфибий. У рыб возникают также парные конечности — грудные и брюшные плавники, на базе которых впоследствии развиваются парные конечности наземных четвероногих животных. Подробнее рассмотрим происхождение и эволюцию парных конечностей. У личинок рыб, а также у современных бесчерепных вдоль тела с обеих сторон тянутся боковые кожные складки, называемые метаплевральными (рис. 14.10). Они не имеют ни скелета, ни собственной мускулатуры, выполняя пассивную роль — стабилизацию положения тела и увеличение площади брюшной поверхности, облегчающие перемещение в водной среде. Вероятно, у предков рыб, переходящих к более активному образу жизни, в этих складках появились мышечные элементы и хрящевые лучи, связанные с сомитами по происхождению и поэтому расположенные метамерно. Такие складки, приобретя подвижность, могут выполнять роль рулей глубины, однако для изменения положения тела в пространстве большее значение имеют их передние и задние отделы, как наиболее удаленные от центра тяжести. Поэтому эволюция шла по пути интенсификации функций крайних отделов и ослабления функций центральных частей.
Рис. 14.10. Формирование передних и задних конечностей из метаплевральных складок: I—III—гипотетические этапы эволюции В результате из передних отделов складок развились грудные, а из задних — брюшные плавники (рис. 14.10). Не исключено, что формированию только двух пар конечностей на боковых сторонах тела предшествовал распад сплошных складок на ряд парных плавников, большее значение из которых также имели передние и задние. Об этом свидетельствует существование ископаемых остатков древнейших низкоорганизованных рыб с многочисленными плавниками (рис. 14.11). За счет слияния оснований хрящевых лучей возникли плечевой и тазовый пояса. Остальные их участки дифференцировались в скелет свободных конечностей.
Рис. 14.11. Древняя акулообразная рыба с многочисленными парными конечностями
У большинства рыб в скелете парных плавников выделяют проксимальный отдел, состоящий из небольшого числа хрящевых или костных пластинок, и дистальный, в состав которого входит большое количество расположенных радиально сегментированных лучей. С поясами конечностей плавники соединены малоподвижно. Они не могут служить опорой телу при передвижении по дну или суше. У кистеперых рыб скелет парных конечностей имеет иное строение. Общее количество их костных элементов уменьшено, и они имеют более крупные размеры. Проксимальный отдел состоит только из одного крупного костного элемента, соответствующего плечевой или бедренной костям передних или задних конечностей. Далее следуют две более мелкие косточки, гомологичные локтевой и лучевой или большой и малой берцовым костям. На них опираются 7—12 радиально расположенных лучей. В соединении с поясами конечностей у такого плавника участвуют только гомологи плечевой или бедренной костей, поэтому плавники кистеперых рыб оказываются активно подвижными (рис. 14.12, А, Б) и могут использоваться не только для изменения направления движения в воде, но и для перемещения по твердому субстрату. Жизнь этих рыб в мелких пересыхающих водоемах в девонском периоде способствовала отбору форм с более развитыми и подвижными конечностями. Наличие у них добавочных органов дыхания (см. разд. 14.3.4) стало второй предпосылкой выхода на сушу и возникновения других адаптации к наземному существованию, результатом чего явилось происхождение земноводных и всей группы Tetrapoda. Первые их представители — стегоцефалы — обладали семи- и пятипалыми конечностями, сохраняющими сходство с плавниками кистеперых рыб (рис. 14.12, B)
Рис. 14.12. Скелет конечности кистеперой рыбы (А), его основание (Б) и скелет передней лапы стегоцефала (В): I—плечевая кость, 2—локтевая кость, 3—лучевая кость
В скелете запястья сохранено правильное радиальное расположение костных элементов в 3—4 ряда, в пястье располагается 7—5 костей, а далее также радиально лежат фаланги 7—5 пальцев. У современных земноводных количество пальцев в конечностях равно пяти или происходит их олигомеризация до четырех. Дальнейшее прогрессивное преобразование конечностей выражается в увеличении степени подвижности соединений костей, в уменьшении количества костей в запястье сначала до трех рядов у амфибий и затем до двух — у пресмыкающихся и млекопитающих. Параллельно уменьшается также и количество фаланг пальцев. Характерно также удлинение проксимальных отделов конечности и укорочение дистальных. Расположение конечностей в ходе эволюции также меняется. Если у рыб грудные плавники находятся на уровне первого позвонка и обращены в стороны, то у наземных позвоночных в результате усложнения ориентации в пространстве появляется шея и возникает подвижность головы, а у пресмыкающихся и особенно у млекопитающих в связи с приподнятием тела над землей передние конечности перемещаются кзади и ориентируются не горизонтально, а вертикально. То же касается и задних конечностей. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|