|
|
Эволюция мочеполовых протоков
В эмбриогенезе всех позвоночных при развитии предпочки вдоль тела, от головного конца к клоаке, закладывается канал, по которому продукты диссимиляции из нефронов поступают во внешнюю среду. Это пронефрический канал. При развитии первичной почки этот канал либо расщепляется на два канала, идущих параллельно, либо второй канал образуется в продольном утолщении стенки первого. Один из них — вольфов — вступает в связь с нефронами первичной почки. Другой — мюллеров — срастается передним концом с одним из нефронов предпочки и образует яйцевод, открывающийся передним концом в целом широкой воронкой, а задним — впадающий в клоаку. Вне зависимости от пола у всех позвоночных обязательно формируются как вольфов, так и мюллеров каналы, однако судьба их различна как у разных полов, так и у представителей разных классов. У самок рыб и земноводных вольфов канал всегда выполняет функцию мочеточника, а мюллеров — яйцевода. У самцов мюллеров канал редуцируется и обе функции — половую и выделительную — выполняет вольфов канал. Семенные канальцы при этом впадают в почку, а сперматозоиды при оплодотворении поступают в воду вместе с мочой. У пресмыкающихся и млекопитающих большая часть вольфова канала не принимает участия в выведении мочи и только его наиболее каудальная часть в области впадения в клоаку образует выпячивание, становящееся мочеточником вторичной почки. Сам же вольфов канал у самцов выполняет функцию семяизвергательного канала. Мюллеров канал у них подвергается редукции. У самок вольфов канал редуцируется (за исключением его каудальной части, формирующей мочеточник), а мюллеров — становится яйцеводом (рис. 14.36). У плацентарных млекопитающих мюллеров канал дифференцируется на собственно яйцевод, матку и влагалище. Будучи парным образованием, как и все элементы половой системы, мюллеров канал сохраняет парность строения у яйцекладущих и частично у сумчатых млекопитающих, у которых имеется два влагалища, две матки и два яйцевода. В дальнейшей эволюции происходит срастание мюллеровых каналов с образованием одного влагалища и матки, которая может быть либо двойной, как у многих грызунов, либо двураздельной, как у хищных; либо двурогой, как у насекомоядных и китообразных, либо простой, как у приматов и человека. Соответственно дифференцировкам мюллерова канала самок у самцов пресмыкающихся и млекопитающих развиваются копулятивные органы. У большинства пресмыкающихся, а также у сумчатых млекопитающих они парные. У плацентарных с одним влагалищем копулятивный орган непарный, но в его развитии обнаруживается срастание парных зачатков.
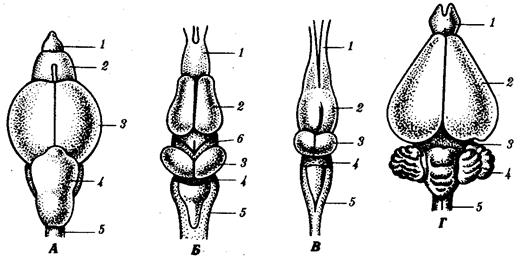
Рис. 14.36. Эволюция почки и мочеполовых каналов. А — нейтральное зародышевое состояние; Б — анамнии; В — амниоты: I—самки, II—самцы; 1—предпочка, 2—первичная почка, 3—канал предпочки, 4—половая железа, 5—мюллеров канал, 6—вольфов канал, 7—мочевой пузырь, 8—клоака, 9—вторичная почка, 10—матка, 11—мочеполовой синус, 12—задняя кишка, 13—половой член, 14—мочеточник вторичной почки, 15—мужская «маточка» В эмбриогенезе человека закладываются парные вольфовы и мюллеровы каналы. Позже в зависимости от пола происходит их редукция. Рудимент мюллерова канала у мужчин располагается в предстательной железе и называется мужской маточкой — utriculus masculinus. Канальцы передней части первичной почки у них вступают в связь с семенниками и преобразуются в придаток семенника — эпидидимис. У плодов женского пола возможно нарушение редукции вольфовых каналов, которые располагаются по бокам от влагалища. Эта аномалия опасна возможностью образования кист и злокачественного перерождения. Распространенными пороками развития являются также различные формы удвоения матки (1 случай на 1000 перинатальных вскрытии). Они развиваются как результат нарушения срастания мюллеровых каналов. Нарушение срастания парных зачатков полового члена в эмбриогенезе человека может привести к формированию такого порока развития, как его удвоение (рис. 14.37).
Рис. 14.37. Аномалии матки и полового члена у человека. А—Д—различные формы удвоения матки и влагалища; Е—удвоение полового члена 101.Филогенез нервной системы позвоночных. Этапы эволюции головного мозга позвоночных. Онтофилогенетические пороки нервной системы у человека. Нервная система хордовых животных, как и у всех многоклеточных, развивается из эктодермы. Она возникла за счет погружения чувствительных клеток, первоначально лежавших на поверхности тела, под его покровы. Это доказывается и сравнительно-анатомическими, и эмбриологическими данными. Действительно, у наиболее примитивного представителя хордовых — ланцетника — центральная нервная система, состоящая из нервной трубки, сохранила функции органа чувств: среди клеток, лежащих внутри нее, имеются отдельные светочувствительные образования — глазки Гессе. Кроме того, основные дистантные органы чувств — зрения, обоняния и слуха — образуются у всех позвоночных первоначально как выпячивания передней части нервной трубки. В эмбриогенезе нервная система формируется вначале всегда в виде полосы утолщенной эктодермы на спинной стороне зародыша, которая впячивается под покровы и замыкается в трубку с полостью внутри — невроцелем. У ланцетника это замыкание еще не полное, поэтому нервная трубка выглядит как желобок (рис. 14.38). Передний конец ее расширен. Он гомологичен головному мозгу позвоночных. Большинство клеток нервной трубки ланцетника не являются нервными, они выполняют опорные или рецепторные функции.
Рис 14.38. Нервная трубка ланцетника: 1 — невроцель, 2 — глазки Гессе У всех позвоночных центральная нервная система является производной нервной трубки, передний конец которой становится головным мозгом, а задний — спинным. Образование головного мозга называют кефализацией. Она связана с усилением двигательной активности позвоночных и необходимостью постоянного анализа раздражении, приходящих из внешней среды, в первую очередь с переднего конца тела. Этот процесс сопровождается также дифференциацией органов чувств, особенно дистантных — обоняния, зрения и слуха. Совместная эволюция органов чувств и головного мозга приводит к возникновению динамических координации между обонятельными рецепторами и передним мозгом, зрительными — и средним, слуховыми — и задним. Видимо, поэтому головной мозг всех современных позвоночных животных в эмбриогенезе закладывается вначале из трех мозговых пузырей — переднего, среднего и заднего — и только позже дифференцируется на пять отделов. Вероятно, предки позвоночных имели более простой головной мозг, развивающийся на основе трех мозговых пузырей. Головной мозг современных взрослых позвоночных всегда состоит из пяти отделов: переднего, промежуточного, среднего, заднего и продолговатого. Внутри головного и спинного мозга расположена общая полость, соответствующая невроцелю. В спинном мозге это спинномозговой канал, а в головном — желудочки мозга. Ткань мозга состоит из серого вещества (скопления нервных клеток) и белого (отростков нервных клеток). Во всех отделах головного мозга различают мантию, располагающуюся над желудочками, и основание, лежащее под ними. В прогрессивной эволюции головного мозга проявляется постепенное усиление роли его передних отделов и мантии по сравнению с задними и основанием. У рыб головной мозг в целом невелик. Слабо развит его передний отдел. Передний мозг не разделен на полушария. Крыша его тонкая, состоит только из эпителиальных клеток и не содержит нервной ткани. Основание переднего мозга включает полосатые тела, от него отходят обонятельные доли. Функционально передний мозг является высшим обонятельным центром. В промежуточном мозге, с которым связаны эпифиз и гипофиз, расположен гипоталамус, являющийся центральным органом эндокринной системы. Средний мозг рыб наиболее развит. Он состоит из двух полушарий и служит высшим зрительным центром. Кроме того, он представляет собой высший интегрирующий отдел головного мозга. Задний мозг содержит мозжечок, осуществляющий регуляцию координации движений. Он развит очень хорошо в связи с перемещением рыб в трехмерном пространстве. Продолговатый мозг обеспечивает связь высших отделов головного мозга со спинным и содержит центры дыхания и кровообращения. Головной мозг такого типа, в котором высшим центром интеграции функций является средний мозг, называют ихтиопсидным. У земноводных головной мозг также ихтиопсидный. Однако передний мозг их имеет большие размеры и разделен на полушария. Крыша его состоит из нервных клеток, отростки которых располагаются на поверхности. Как и у рыб, больших размеров достигает средний мозг, также представляющий собой высший интегрирующий центр и центр зрения. Мозжечок несколько редуцирован в связи с примитивным характером движений. Условия наземного существования пресмыкающихся требуют более сложной морфофункциональной организации мозга. Передний мозг — наиболее крупный отдел по сравнению с остальными. В нем особенно развиты полосатые тела. К ним переходят функции высшего интегративного центра. На поверхности крыши впервые появляются островки коры очень примитивного строения, ее называют древней — archicortex. Средний мозг теряет значение ведущего отдела, и относительные размеры его сокращаются. Мозжечок сильно развит благодаря сложности и многообразию движений пресмыкающихся. Головной мозг такого типа, в котором ведущий отдел представлен полосатыми телами переднего мозга, называют зауропсидным. У млекопитающих — маммалийный тип мозга. Для него характерно сильное развитие переднего мозга за счет коры, которая развивается на основе небольшого островка коры пресмыкающихся и становится интегрирующим центром мозга. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называется новой корой — neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участки. Характерным является также наличие комиссуры между обоими полушариями, в которой располагаются волокна, связывающие их воедино. Промежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Два передних связаны со зрительным анализатором, два задних—со слуховым. Очень хорошо развит мозжечок (рис. 14.39).
Рис. 14.39. Эволюция головного мозга позвоночных. А—рыба; Б—земноводное; В—пресмыкающееся; Г—млекопитающее: 1—обонятельные доли, 2—передний мозг, 3—средний мозг, 4—мозжечок, 5—продолговатый мозг, 6—промежуточный мозг
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференцировка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца. Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити — filum terminale. Нервы, идущие к заднему концу тела, проходят по позвоночному каналу самостоятельно, образуя так называемый конский хвост — cauda equina. Основные этапы эволюции центральной нервной системы отражаются и в онтогенезе человека. На стадии нейруляции закладывается нервная пластинка, превращающаяся в желобок и затем в трубку. Передний конец трубки образует сначала три мозговых пузыря (рис. 14.40): передний (I), средний (II) и задний (III). Вслед за этим передний пузырь подразделяется на два, дифференцирующихся на передний (7) и промежуточный (2) мозг — telencephalon, diencephalon. Средний мозговой пузырь развивается в средний (3) мозг — mesencephalon, a задний — в задний (4) мозг — metencephalon — и продолговатый (5) мозг — medulla oblongata.
Рис. 14.40. Мозговые пузыри в эмбриогенезе мозга человека. А — стадия трех мозговых пузырей; Б — стадия пяти мозговых пузырей (остальные пояснения см. в тексте)
Задний конец спинного мозга редуцируется, превращаясь в терминальную нить. Позже скорости роста спинного мозга и позвоночника оказываются разными, и к моменту рождения конец спинного мозга оказывается на уровне третьего, а у взрослого человека — уже на уровне первого поясничного позвонка. Головной мозг рано начинает развиваться по пути, характерному для млекопитающих и человека. Первично почти прямая нервная трубка резко изгибается в области будущего продолговатого и среднего мозга. На этом фоне большие полушария переднего мозга растут с особенно большой скоростью. В результате головной мозг оказывается расположенным над лицевым черепом. Дифференцировка коры приводит к развитию извилин, борозд и формированию высших сенсорных и двигательных центров, в том числе центров письменной и устной речи и др., характерных только для человека (см. § 15.3). Центральная нервная система столь важна для интеграции индивидуального развития человека, что большинство ее врожденных пороков несовместимы с жизнью. Среди пороков спинного мозга, онтогенетические механизмы которых известны, отметим рахисхиз,нли платиневрию,— отсутствие замыкания нервной трубки (рис. 14.41). Эта аномалия связана с нарушением клеточных перемещений и адгезии в зоне формирования нервной трубки в процессе нейруляции. Аномалия переднего мозга — прозэнцефалия — выражается в нарушении морфо-генеза мозга, при котором полушария оказываются неразделенными, а кора — недоразвита. Этот порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью. Часто встречается у мертворожденных при различных хромосомных и генных синдромах. Нарушения дифференцировки коры — агирия (отсутствие извилин) и олигогирия с пахигирией (малое количество утолщенных извилин) — сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей умирают в течение первого года жизни.
Рис. 14.41. Рахисхиз (платиневрия) 102.Филогенез эндокринной системы. Гормоны. Эволюционные преобразования желез внутренней секреции у хордовых животных. Онтофилогенетические пороки эндокринной системы у человека. Эндокринная система наряду с нервной является ведущим аппаратом интеграции многоклеточного организма, обеспечивая гуморальную регуляцию функций органов. Эта регуляция осуществляется гормонами — биологически активными веществами разной химической природы, выделяемыми железами внутренней секреции. Действие гормонов строго специфично: разные гормоны действуют на разные органы, вызывая определенные изменения их функционирования. Железы внутренней секреции не имеют протоков и выделяют гормоны непосредственно в кровь, что облегчает их транспорт к органам-мишеням. Клетки органов-мишеней на своих мембранах имеют специфические рецепторы, с которыми связываются гормоны, вызывая определенные изменения их метаболизма. Гуморальная регуляция эволюционно возникла значительно раньше нервной потому, что она более проста и не требует развития таких сложных структур, как нервная система. Обсуждая вопрос о филогенезе эндокринной системы хордовых, необходимо рассмотреть происхождение и эволюцию как гормонов, так и самих желез внутренней секреции. Гормоны
Химическая структура гормонов многообразна. Это могут быть белки (инсулин, пептидные нейрогормоны), стероиды (половые гормоны), продукты метаболизма отдельных аминокислот (тироксин, адреналин). Они могут иметь и другое строение. Это свидетельствует о том, что гормоны разнообразны и по происхождению. Главное, что их объединяет, — способность специфически изменять клеточный метаболизм при контакте с цитоплазматической мембраной. Предполагают, что уже древние одноклеточные организмы использовали биологически активные вещества для межклеточных коммуникаций. Некоторые вещества такого рода, обладающие регуляторными функциями, могли действовать как на отдельные клетки простейших, так позже и на клетки многоклеточных организмов. Впоследствии они и стали выполнять функции гормонов. Интересно, что в прогрессивной эволюции гуморальной регуляции структура самих гормонов может и не меняться. Доказательством этого является обнаружение таких известных гормонов, как адреналин, норадреналин и некоторые другие, в клетках простейших и низших растений, где они выполняют функции регуляторов клеточного деления, движения ресничек и вакуолей. В эмбриогенезе многоклеточных ряд гормонов выявляется уже в первые часы и дни развития. В процессе дробления они регулируют течение клеточного цикла. Позже — перемещения клеток и образование межклеточных контактов, действуя либо внутри клеток, их продуцирующих, либо на близлежащие клетки. Гормоны приобретают свойства дистантных регуляторов в филогенезе только у трехслойных животных, а в онтогенезе многоклеточных — соответственно на стадии первичного органогенеза. При неизменности химической структуры функции гормонов могут нередко изменяться. Так, гормон пролактин, выделяющийся у млекопитающих и человека гипофизом и регулирующий секрецию молока млечными железами, обнаружен также у рыб, земноводных и птиц. У первых он регулирует выделение кожными железами слизи, которой питаются мальки, у вторых — образование оболочек икринок в яйцеводах, у третьих — некоторые элементы брачного поведения, а также выделение у кормящих родителей «зобного молочка». Как видно из этого примера, эволюция действия гормонов может быть канализированной выполнением одной и той же функции в самом широком смысле. Действительно, все перечисленные функции пролактина у животных, находящихся на разных уровнях организации, имеют отношение к обеспечению успешности размножения. Функции других гормонов, например адреналина, могут в филогенетическом ряду тех же позвоночных практически не меняться, обеспечивая у всех регуляцию в первую очередь энергетического обмена. При этом часто гормоны продолжают выполнять те функции, которые первично проявлялись еще у одноклеточных и низших многоклеточных животных. Так, адреналин в организме млекопитающих и человека замедляет прохождение клеток по клеточному циклу и вступление их в митоз. Из главной эта функция у высокоорганизованных животных превратилась во второстепенную. Таким образом, на филогенез гормонов, так же как и на эволюцию органов, распространяются основные закономерности макроэволюции (см. гл. 13). 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|