
|
|
Гладкая мышечная ткань нейрального происхожденияМиоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы — суживающую и расширяющую зрачок. 37. Морфо - функциональная характеристика гладкого миоцита ( СМ и ЭМ). Структурно-функциональной единицей гладкой, или неисчерченной, мышечной ткани является гладко-мышечная клетка, или гладкий миоцит — это веретеновидная клетка длиной 20—500 мкм, шириной 5—8 мкм. Ядро клетки палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается. Органеллы общего значения, среди которых много митохондрий, сосредоточены в цитоплазме около полюсов ядра. Аппарат Гольджи и гранулярная эндо плазматическая сеть развиты слабо, что свидетельствует о малой активности синтетических функций. Рибосомы в большинстве своем расположены свободно. Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерий обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм.
38. Особенности строения сократительного и опорного аппарата гладкого миоцита. Актиновые и миозиновые филаменты. Механизм мышечного сокращения.
39 . Нервная ткань. Гистогенез. Производные нервной трубки (нейробласты, глиобласты), нервного гребня и нейральных плакод. НЕРВНАЯ ТКАНЬ Часть первая – общая характеристика, классификация и развитие нервной ткани. Нервная ткань — это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и его передачи. Она является основой строения органов нервной системы, обеспечивающих регуляцию всех тканей и органов, их интеграцию в организме и связь с окружающей средой. В нервной ткани выделяют два типа клеток – нервные и глиальные. Нервные клетки(нейроны, или нейроциты) — основные структурные компоненты нервной ткани, выполняющие специфическую функцию. Нейроглия обеспечивает существование и функционирование нервных клеток, осуществляя опорную, трофическую, разграничительную, секреторную и защитную функции. Развитие
А — стадия нервной пластинки; Б - стадия нервного желобка; В - стадия нервной трубки. Нервная ткань развивается из дорсальной эктодермы. У 18-дневного эмбриона человека эктодерма по средней линии спины дифференцируется и утолщается, формируя нервную пластинку, латеральные края которой приподнимаются, образуя нервные валики, а между валиками формируется нервный желобок. Передний конец нервной пластинки расширяется, образуя позднее головной мозг. Латеральные края продолжают подниматься и растут медиально, пока не встретятся и не сольются по средней линии в нервную трубку, которая отделяется от лежащей над ней кожной эктодермы. Полость нервной трубки сохраняется у взрослых в виде системы желудочков головного мозга и центрального канала спинного мозга. Часть клеток нервной пластинки не входит в состав ни нервной трубки, ни кожной эктодермы, а образует скопления по бокам от нервной трубки, которые сливаются в рыхлый тяж, располагающийся между нервной трубкой и кожной эктодермой, — это нервный гребень(или ганглиозная пластинка). Из нервной трубки в дальнейшем формируются нейроны и макроглия центральной нервной системы. Нервный гребень дает начало нейронам чувствительных и автономных ганглиев, клеткам мягкой мозговой и паутинной оболочек мозга и некоторым видам глии: нейролеммоцитам (шванновским клеткам), клеткам-сателлитам ганглиев. Из нервного гребня развиваются также клетки мозгового вещества надпочечников, меланоциты кожи, часть клеток APUD-системы, сенсорные клетки каротидных телец. В формировании ганглиев V, VII, IX и X пар черепных нервов принимают участие, кроме нервного гребня, также нейрогенные плакоды, представляющие собой утолщения эктодермы по бокам формирующейся нервной трубки в краниальном отделе зародыша. Нервная трубка на ранних стадиях эмбриогенеза представляет собой многорядный нейроэпителий, состоящий из вентрикулярных, или нейроэпителиальных клеток. В дальнейшем в нервной трубке дифференцируется 4 концентрических зоны: · внутренняя - вентрикулярная (или эпендимная) зона, · вокруг нее – субвентрикулярная зона, · затем промежуточная (или плащевая, или же мантийная, зона) и, наконец, · наружная - краевая (или маргинальная) зона нервной трубки. Вентрикулярная (эпендимная), внутренняя, зона состоит из делящихся клеток цилиндрической формы. Вентрикулярные (или матричные) клетки являются предшественниками нейронов и клеток макроглии. Субвентрикулярная зона состоит из клеток, сохраняющих высокую пролиферативную активность и являющихся потомками матричных клеток. Промежуточная (плащевая, или мантийная) зона состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон — нейробластов и глиобластов. Нейробласты утрачивают способность к делению и в дальнейшем дифференцируются в нейроны. Глиобласты продолжают делиться и дают начало астроцитам и олигодендроцитам. Способность к делению не утрачивают полностью и зрелые глиоциты. Новообразование нейронов прекращается в раннем постнатальном периоде.
Поскольку число нейронов в головном мозге составляет примерно 1 триллион, очевидно, в среднем в течение всего пренатального периода в 1 мин формируется 2,5 миллиона нейронов. Из клеток плащевого слоя образуются серое вещество спинного и часть серого вещества головного мозга. Маргинальная зона (или краевая вуаль) формируется из врастающих в нее аксонов нейробластов и макроглии и дает начало белому веществу. В некоторых областях головного мозга клетки плащевого слоя мигрируют дальше, образуя кортикальные пластинки — скопления клеток, из которых формируется кора большого мозга и мозжечка (т.е. серое вещество). По мере дифференцировки нейробласта, изменяется субмикроскопическое строение его ядра и цитоплазмы. Специфическим признаком начавшейся специализации нервных клеток следует считать появление в их цитоплазме тонких фибрилл — пучков нейрофиламентов и микротрубочек. Количество нейрофиламентов, содержащих белок — нейрофиламентный триплет, в процессе специализации увеличивается. Тело нейробласта постепенно приобретает грушевидную форму, а от его заостренного конца начинает развиваться отросток — аксон. Позднее дифференцируются другие отростки — дендриты. Нейробласты превращаются в зрелые нервные клетки — нейроны. Между нейронами устанавливаются контакты (синапсы). В процессе дифференцировки нейронов из нейробластов различают до-медиаторный и медиаторный периоды. Для домедиаторного периода характерно постепенное развитие в теле нейробласта органелл синтеза — свободных рибосом, а затем эндоплазматической сети. В медиаторном периоде у юных нейронов появляются первые пузырьки, содержащие нейромедиатор, а в дифференцирующихся и зрелых нейронах отмечаются: значительное развитие органелл синтеза и секреции, накопление медиаторов и поступление их в аксон, образование синапсов. Несмотря на то, что формирование нервной системы завершается только в первые годы после рождения, известная пластичность центральной нервной системы сохраняется до старости. Эта пластичность может выражаться в появлении новых терминалей и новых синаптических связей. Нейроны центральной нервной системы млекопитающих способны формировать новые ветви и новые синапсы. Пластичность проявляется в наибольшей степени в первые годы после рождения, но частично сохраняется и у взрослых — при изменении уровней гормонов, обучении новым навыкам, травме и других воздействиях. Хотя нейроны постоянны, их синаптические связи могут модифицироваться в течение всей жизни, что может выражаться, в частности, в увеличении или уменьшении их числа. Пластичность при малых повреждениях мозга проявляется в частичном восстановлении функций. В популяции нейронов, начиная с ранних стадий развития нервной системы и в течение всего онтогенеза, имеет место массовая гибель клеток. Эта запрограммированная физиологическая гибель клеток наблюдается как в центральной, так и в периферической нервной системе. У человека ежегодно погибает около 10 млн нервных клеток. 40.Морфологическая и функциональная классификация нейронов. Морфофункциональные зоны нейрона (классификация по Бодиану). Органеллы общего и специального значения. Существует множество классификаций нервных клеток, так как нейроны отличаются по размерам и форме перикариона, числу отростков, их синаптическим связям, характеру ветвления дендритов, электрофизиологическим характеристикам, химии нейромедиаторов, позиции в нейронных сетях и множеству других характеристик. В зависимости от класса выполняемой функции выделяют афферентные (чувствительные, сенсорные), эфферентные (двигательные, моторные) и вставочные нервные клетки (ассоциативные нейроны, или интернейроны). Афферентные нейроны проводят возбуждение от рецепторов периферических органов в структуры ЦНС. Эфферентные нейроны осуществляют передачу сигналов от ЦНС к органам-эффекторам (мышцам и железам). Ассоциативные нейроны проводят возбуждение между нейронами. Морфологическая классификация. По количеству отростков нейроны разделяют на псевдоуниполярные, биполярные и мультиполярные. Псевдоуниполярные нейроны имеют один короткий отросток, который разделяется на некотором расстоянии от сомы на два длинных — дендрит и аксон. К псевдоуниполярным относятся нейроны сенсорных ганглиев спинного мозга. Биполярные нейроны имеют один дендрит и один аксон. Этот вид нейронов встречается в периферическом отделе зрительного, обонятельного и слухового анализаторов. Мультиполярные нейроны имеют один аксон и несколько дендритов, это наиболее распространённый вид нейронов. К ним относятся мотонейроны спинного мозга. Исходя из характера электрогенеза в различных зонах нервных клеток (рецептивная и интегративная зоны, начальный сегмент аксона, аксон, терминали аксона), предложена универсальная классификация (рис. 5–2, см. также рис. 5–1). Эта классификация учитывает направление (вход и выход) сигналов в пределах одной нервной клетки, но находящейся в составе нейронных сетей или цепочек, а также характер мембранного электрогенеза (изменения МП в рецептивной и пресинаптической зонах и передача ПД по аксону).
Рис. 5–2. Универсальная классификация частей различных типов нейронов [11]. Дендриты — рецепторная зона — возбуждающий или тормозящий вход. Перикарион с дендритами — интегративная зона. Аксон — проводящая возбуждение (ПД) часть нейрона, его концевые разветвления образуют синапсы (выход) с соседними клетками. А–Г — афферентные (чувствительные) нейроны; Д — мотонейрон; Е — интернейроны. Специфическими органеллами нейрона являются нейро-филаменты и нейротубулы+ -тигровидное вещество, или вещество Ниссля, которое представляет собой участки цитоплазмы с большим содержанием рибосом.. Нейрофиламенты представляют собой промежуточные филаменты диаметром 8-10 нм, образованные фибриллярными белками (белками так называемого нейрофибриллярного триплета, или нейрофибрилляр-ными кислыми белками). Основными функциями данной органеллы являются опорно-каркасная, обеспечение стабильной формы нейрона и нервной системы в целом. Аналогичную роль играют тонкие микрофи-ламенты (поперечный диаметр 6-8 нм), содержащие белки актины. В отличие от подобных микрофиламентов в других тканях и клетках, они не соединяются с микромиозинами, что делает невозможным активные сократительные функции в зрелых нервных клетках. Нейротубулы по основным принципам своего строения практически не отличаются от микротрубочек. Они, как и все микротрубочки, имеют поперечный диаметр около 24 нм и на поперечном разрезе сформированы 13 молекулами глобулярных белков тубулинов. Как и везде, они поляризованы. В отличие от большинства микротрубочек в других клетках, нейротубулы весьма стабильны. Тубулин в них находится в метилированной форме и нередко кэпирован (концы нейроту-бул прикрыты белковыми молекулами, функция которых заключается в стабилизации нейротубул и предохранении их от разрушения). В нервной ткани они выполняют очень важную, если не сказать, уникальную роль. Они несут опорно-каркасную функцию, обеспечивают процессы циклоза, направляя органеллы и включения. Полярность ор-ганеллы, в которой имеется отрицательно и положительно заряженный конец, позволяет контролировать диффузионно-транспортные потоки в аксоне (так называемый быстрый и медленный аксоток). Кроме того, значительное число нейрофизиологов приписывает микротрубочкам роль хранилища поступающей в мозг информации. 41.Транспортные процессы в нейроне. Дендритный и аксонный транспорт Роль плазмолеммы нейронов в рецепции, генерации и проведении нервного импульса. Аксональный (точнее аксоплазматический) транспорт — это перемещение веществ от тела в отростки и от отростков в тело нейрона. Он направляется нейротубулами, а в транспорте участвуют белки — кинезин и динеин. Транспорт веществ от тела клетки в отростки называется прямым, или антероградным, транспорт веществ от отростков к телу — обратным, или ретроградным. Аксональный транспорт представлен двумя главными компонентами: быстрым компонентом (400—2000 мм в сутки) и медленным (1—2 мм в сутки). Обе транспортные системы присутствуют как в аксонах, так и в дендритах. Антероградная быстрая система проводит мембранные структуры, включая компоненты мембраны, митохондрии, пузырьки, содержащие пептиды, предшественники нейромедиаторов и другие белки. Ретроградная быстрая система проводит использованные материалы для деградации в лизосомах, распределения и рециркуляции и, возможно, факторы роста нервов. Нейротубулы — органеллы, ответственные за быстрый транспорт, который называется также нейротубулозависимым. Каждая нейротубула содержит несколько путей, вдоль которых движутся различные частички. АТФ и ионы Са2+ обеспечивают эти движения. На одной микротубуле пузырьки могут обгонять другие пузырьки, движущиеся в том же направлении. Два пузырька могут двигаться в противоположных направлениях одновременно по различным путям одной нейротубулы. Медленный транспорт — это антероградная система, проводящая белки и другие вещества для обновления и поддержания аксоплазмы зрелых нейронов и обеспечения аксоплазмой роста аксонов и дендритов при развитии и регенерации. Во ещё раз по нормальному Аксональный транспорт – это непрерывная циркуляция аксоплазмы от перикариона (антеградный ток) и назад к нему (ретроградный ток). Антеградный ток: а) медленный поток: 0,1 – 3 мм/сутки, который несёт новосинтезированную аксоплазму к окончанию аксона (глиальная оболочка выполняет при это роль перистальтики); б) быстрый поток: 100-500 мм/сутки, а в нейросекреторных клетках гипоталамуса – 2800мм/сутки. Этот поток несёт вещества для синаптической функции: ферменты, гликопротеиды, фосфолипиды, митохондрии. Ретроградный ток происходит от дистальной части аксона в сторону перикариона и осуществляется со скоростью быстрого потока. Это перемешение белков и других веществ, захваченных нервными окончаниями. Дендритный транспорт осуществляет перенос белков и ферментов (ацетилхолинестераза) от тела к дендритам со скоростью 3мм/сутки (медленный поток). Механизм транспорта. Транспорт обеспечивается микротрубочками и связанными с ними белками кинезинами и динеинами и происходит с затратой АТФ. Кинезин отвечает за антеградный транспорт, а динеин – за ретроградный транспорт, который происходит по поверхности микротрубочек, а не внутри них. Трансверсальный транспорт – это передача кислорода, энергетических субстратов,удаление продуктов метаболизма через местное кровеносное русло в области перехватов Ранвье. После прекращения кровоснабжения нервное волокно теряет способность к проведению возбуждения.
Плазмолемма нейрона является возбудимой мембраной, т.е. обладает способностью генерировать и проводить импульс. Ее интегральными белками являются белки, функционирующие как ионно-избирательные каналы, и рецепторные белки, вызывающие реакции нейронов на специфические стимулы. В нейроне мембранный потенциал покоя равен —60 —70 мВ. Потенциал покоя создается за счет выведения Na+ из клетки. Большинство Na+- и К+-каналов при этом закрыты. Переход каналов из закрытого состояния в открытое регулируется мембранным потенциалом. В результате поступления возбуждающего импульса на плазмолемме клетки происходит частичная деполяризация. Когда она достигает критического (порогового) уровня, натриевые каналы открываются, позволяя ионам Na+ войти в клетку. Деполяризация усиливается, и при этом открывается еще больше натриевых каналов. Калиевые каналы также открываются, но медленнее и на более продолжительный срок, что позволяет К+ выйти из клетки и восстановить потенциал до прежнего уровня. Через 1—2 мс (т.н. рефрактерный период) каналы возвращаются в нормальное состояние, и мембрана может вновь отвечать на стимулы. Итак, распространение потенциала действия обусловлено вхождением в нейрон ионов Na+, которые могут деполяризовать соседний участок плазмолеммы, что в свою очередь создает потенциал действия на новом месте. При окрашивании нервной ткани анилиновыми красителями в цитоплазме нейронов выявляется хроматофильная субстанция в виде базофильных глыбок и зерен различных размеров и форм (другие названия хроматофильной субстанции - тигроид, тельца Ниссля). Базофильные глыбки локализуются в перикарионах и дендритах нейронов, но никогда не обнаруживаются в аксонах и их конусовидных основаниях — аксональных холмиках. Базофилия глыбок объясняется высоким содержанием рибонуклеопротеидов. Каждая глыбка хроматофильной субстанции состоит из цистерн гранулярной эндоплазматической сети, свободных рибосом и полисом. Для поддержания целостности нейронов и выполнения ими функций нейронам требуется огромное количество белков. Для аксонов, не имеющих органелл белкового синтеза, характерен постоянный ток цитоплазмы от перикариона к терминалям со скоростью 1—3 мм в сутки. 42.Нейроглия. Морфофункциональная характеристика . Локализация. Строение. Функция. Нейроглия Нейроны — это высокоспециализированные клетки, существующие и функционирующие в строго определенной среде. Такую среду им обеспечивает нейроглия. Нейроглия выполняет следующие функции: опорную, трофическую, разграничительную, поддержание постоянства среды вокруг нейронов, защитную, секреторную. Различают глию центральной и периферической нервной системы. Клетки глии центральной нервной системы делятся на макроглию и микроглию. Макроглия Макроглия развивается из глиобластов нервной трубки и включает: эпендимоциты, астроциты и олигодендроглиоциты.
Протоплазматические астроциты характеризуются короткими сильно ветвящимися отростками и светлым сферическим ядром. Отростки астроцитов тянутся к базальным мембранам капилляров, к телам и дендритам нейронов, окружая синапсы и отделяя (изолируя) их друг от друга, а также к мягкой мозговой оболочке, образуя пиоглиальную мембрану, граничащую с субарахноидальным пространством. Подходя к капиллярам, их отростки образуют расширенные «ножки», полностью окружающие сосуд. Астроциты накапливают и передают вещества от капилляров к нейронам, захватывают избыток экстрацеллюлярного калия и других веществ, таких как нейромедиаторы, из экстрацеллюлярного пространства после интенсивной нейрональной активности.
Микроглия Микроглия представляет собой фагоцитирующие клетки, относящиеся к системе мононуклеарных фагоцитов и происходящие из стволовой кроветворной клетки (возможно, из премоноцитов красного костного мозга). Функция микроглии — защита от инфекции и повреждения, и удаление продуктов разрушения нервной ткани. Клетки микроглии характеризуются небольшими размерами, телами продолговатой формы. Их короткие отростки имеют на своей поверхности вторичные и третичные ответвления, что придает клеткам «колючий» вид. Описанная морфология характерна для типичной (ветвистой, или покоящейся) микроглии полностью сформированной центральной нервной системы. Она обладает слабой фагоцитарной активностью. Ветвистая микроглия встречается как в сером, так и в белом веществе центральной нервной системы. В развивающемся мозгу млекопитающих обнаруживается временная форма микроглии — амебоидная микроглия. Клетки амебоидной микроглии формируют выросты – филоподии и складки плазмолеммы. В их цитоплазме присутствуют многочисленные фаголизосомы и пластинчатые тельца. Амебоидные микроглиальные тельца отличаются высокой активностью лизосомальных ферментов. Активно фагоцитирующая амебоидная микроглия необходима в раннем постнатальном периоде, когда гематоэнцефалический барьер еще не вполне развит и вещества из крови легко попадают в центральную нервную систему. Считают также, что она способствует удалению обломков клеток, появляющихся в результате запрограммированной гибели избыточных нейронов и их отростков в процессе дифференцировки нервной системы. Полагают, что, созревая, амебоидные микроглиальные клетки превращаются в ветвистую микроглию. Реактивная микроглия появляется после травмы в любой области мозга. Она не имеет ветвящихся отростков, как покоящаяся микроглия, не имеет псевдоподий и филоподий, как амебоидная микроглия. В цитоплазме клеток реактивной микроглии присутствуют плотные тельца, липидные включения, лизосомы. Есть данные о том, что реактивная микроглия формируется вследствие активации покоящейся микроглии при травмах центральной нервной системы. Рассмотренные выше глиальные элементы относились к центральной нервной системе. Глия периферической нервной системы в отличие от макроглии центральной нервной системы происходит из нервного гребня. К периферической нейроглии относятся: нейролеммоциты (или шванновские клетки) и глиоциты ганглиев (или мантийные глиоциты). Нейролеммоциты Шванна формируют оболочки отростков нервных клеток в нервных волокнах периферической нервной системы. Мантийные глиоциты ганглиев окружают тела нейронов в нервных узлах и участвуют в обмене веществ этих нейронов. 43.Безмиелиновые и миелиновые нервные волокна. Ультраструктурная организация миелинового нервного волокна. Нервные волокна Отростки нервных клеток, покрытые оболочками, называются нервными волокнами. По строению оболочек различают миелиновые и безмиелиновые нервные волокна. Отросток нервной клетки в нервном волокне называют осевым цилиндром, или аксоном, так как чаще всего (за исключением чувствительных нервов) в составе нервных волокон находятся именно аксоны.
Безмиелиновые нервные волокна находятся преимущественно в составе автономной, или вегетативной, нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в соседнее. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. По мере погружения осевых цилиндров в тяж нейролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану —мезаксон, на которой как бы подвешен осевой цилиндр. Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Они также состоят из осевого цилиндра, «одетого» оболочкой из нейролеммоцитов Шванна, но диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее.
В процессе миелинизации аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита —мезаксон. Мезаксон удлиняется, концентрически наслаивается (как бы накручивается) на осевой цилиндр и образует вокруг него плотную слоистую зону — миелиновый слой. Отсутствие миелинового слоя в области узловых перехватов объясняется тем, что в этом участке волокна кончается один нейролеммоцит и начинается другой. Осевой цилиндр в этом месте частично прикрыт интердигитирующими отростками нейролеммоцитов. Оболочка аксона (аксолемма) обладает в области перехвата значительной электронной плотностью.
Миелиновые волокна центральной нервной системы не имеют насечек миелина, а нервные волокна не окружены базальными мембранами. 44. Механизм и скорость проведения нервного импульса по нервным волокнам ( потенциал покоя, действия, реполяризация ). Регенерация нервных волокон. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
 Нейруляция (схема).
Нейруляция (схема).
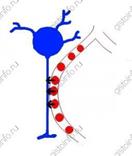
 Эпендимоциты выстилают желудочки головного мозга и центральный канал спинного мозга. Эти клетки цилиндрической формы. Они образуют слой типа эпителия, носящий название эпендимы. Между соседними клетками эпендимы имеются щелевидные соединения и пояски сцепления, но плотные соединения отсутствуют, так что цереброспинальная жидкость может проникать между эпендимоцитами в нервную ткань. Большинство эпендимоцитов имеют подвижные реснички, вызывающие ток цереброспинальной жидкости. Базальная поверхность большинства эпендимоцитов ровная, но некоторые клетки имеют длинный отросток, идущий глубоко в нервную ткань. Такие клетки называются таницитами. Они многочисленны в дне III желудочка. Считается, что эти клетки передают информацию о составе цереброспинальной жидкости на первичную капиллярную сеть воротной системы гипофиза. Эпендимный эпителий сосудистых сплетений желудочков продуцируетцереброспинальную жидкость (ликвор).
Эпендимоциты выстилают желудочки головного мозга и центральный канал спинного мозга. Эти клетки цилиндрической формы. Они образуют слой типа эпителия, носящий название эпендимы. Между соседними клетками эпендимы имеются щелевидные соединения и пояски сцепления, но плотные соединения отсутствуют, так что цереброспинальная жидкость может проникать между эпендимоцитами в нервную ткань. Большинство эпендимоцитов имеют подвижные реснички, вызывающие ток цереброспинальной жидкости. Базальная поверхность большинства эпендимоцитов ровная, но некоторые клетки имеют длинный отросток, идущий глубоко в нервную ткань. Такие клетки называются таницитами. Они многочисленны в дне III желудочка. Считается, что эти клетки передают информацию о составе цереброспинальной жидкости на первичную капиллярную сеть воротной системы гипофиза. Эпендимный эпителий сосудистых сплетений желудочков продуцируетцереброспинальную жидкость (ликвор). Астроциты — клетки отростчатой формы, бедные органеллами. Они выполняют в основном опорную и трофическую функции. Различают два типа астроцитов - протоплазматические и волокнистые. Протоплазматические астроциты локализуются в сером веществе центральной нервной системы, а волокнистые астроциты - преимущественно в белом веществе.
Астроциты — клетки отростчатой формы, бедные органеллами. Они выполняют в основном опорную и трофическую функции. Различают два типа астроцитов - протоплазматические и волокнистые. Протоплазматические астроциты локализуются в сером веществе центральной нервной системы, а волокнистые астроциты - преимущественно в белом веществе. Олигодендроциты – имеют более мелкие по сравнению с астроцитами и более интенсивно окрашивающиеся ядра. Их отростки немногочисленны. Олигодендроглиоциты присутствуют как в сером, так и в белом веществе. В сером веществе они локализуются вблизи перикарионов. В белом веществе их отростки образуют миелиновый слой в миелиновых нервных волокнах, причем, в противоположность аналогичным клеткам периферической нервной системы – нейролеммоцитам, один олигодендроглиоцит может участвовать в миелинизации сразу нескольких аксонов.
Олигодендроциты – имеют более мелкие по сравнению с астроцитами и более интенсивно окрашивающиеся ядра. Их отростки немногочисленны. Олигодендроглиоциты присутствуют как в сером, так и в белом веществе. В сером веществе они локализуются вблизи перикарионов. В белом веществе их отростки образуют миелиновый слой в миелиновых нервных волокнах, причем, в противоположность аналогичным клеткам периферической нервной системы – нейролеммоцитам, один олигодендроглиоцит может участвовать в миелинизации сразу нескольких аксонов. В центральной нервной системе оболочки отростков нейронов образуются отростками олигодендроглиоцитов, а в периферической —нейролеммоцитами Шванна.
В центральной нервной системе оболочки отростков нейронов образуются отростками олигодендроглиоцитов, а в периферической —нейролеммоцитами Шванна. Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии—насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы (1—2 мм) видны участки волокна, лишенные миелинового слоя, — это т.н. узловатые перехваты, или перехваты Ранвье.
Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии—насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы (1—2 мм) видны участки волокна, лишенные миелинового слоя, — это т.н. узловатые перехваты, или перехваты Ранвье. Отрезок волокна между смежными перехватами называется межузловым сегментом. Длина межузлового сегмента, так же как и толщина миелинового слоя, зависит от толщины осевого цилиндра. Насечка миелина (Шмидта—Лантермана) представляет собой участок миелинового слоя, где завитки мезаксона лежат неплотно друг к другу, образуя спиральный туннель, идущий снаружи внутрь и заполненный цитоплазмой нейролеммоцита, т.е. место расслоения миелина. Снаружи от нейролеммоцита располагается базальная мембрана.
Отрезок волокна между смежными перехватами называется межузловым сегментом. Длина межузлового сегмента, так же как и толщина миелинового слоя, зависит от толщины осевого цилиндра. Насечка миелина (Шмидта—Лантермана) представляет собой участок миелинового слоя, где завитки мезаксона лежат неплотно друг к другу, образуя спиральный туннель, идущий снаружи внутрь и заполненный цитоплазмой нейролеммоцита, т.е. место расслоения миелина. Снаружи от нейролеммоцита располагается базальная мембрана.