
|
|
НЕЙРОМОДЕЛЬНЫЙ ПОДХОД К ПОСТРОЕНИЮ ИНТЕЛЛЕКТУАЛЬНЫХ СИСТЕМ[П.Д. Басалин, К.В. Безрук, М.В. Радаева. МОДЕЛИ И МЕТОДЫ ИНТЕЛЛЕКТУАЛЬНОЙ ПОДДЕРЖКИ ПРОЦЕССОВ ПРИНЯТИЯ РЕШЕНИЙ. Учебное пособие. Нижегородский государственный университет им. Н.И. Лобачевского.Национальный исследовательский университет.-Нижний Новгород-
7.1. Биологические основы нейромодельного подхода
Первые исследования по анатомической организации мозга связаны с работами испанского гистолога Сантьяго Рамон-и-Кахаля, датируемыми 1911 годом. Именно он, используя предложенный итальянским гистологом Камилло Гольджи метод окраски нейронов (нервных клеток) солями серебра (серебро избирательно проникает только в нейроны), выявил клеточную (нейронную) структуру мозга. Нейроны (термин предложил Вильгельм фон Вальдейер) представляют собой возбудимые клетки, являющиеся основными функциональными и структурными единицами центральной нервной системы. Они принимают поступающие на них из внешней среды или от других возбудимых клеток сигналы (возбуждения) и, в зависимости от интенсивности этих воздействий, генерируют нервные импульсы, которые передают на другие нейроны, мышцы, железы. Именно эти клетки обеспечивают способность мозга воспринимать и обрабатывать информацию. Головной мозг человека содержит порядка 20 млрд нейронов и 300 млрд межнейронных соединений. Он состоит из левого и правого полушарий, на поверхности которых находится тонкий слой серого вещества. Это кора головного мозга (кортекс), состоящая в основном из нейронов. Кортекс ответствен за интеллектуальные, творческие процессы (мышление), протекающие в человеческом мозге. Левое полушарие мозга реализует аналитический способ обработки информации, суть которого состоит в разложении познаваемого целого на элементарные составляющие известной природы. Оно работает подобно цифровому компьютеру, реализуя линейный (последовательный) способ действий над буквенно-цифровой информацией. При этом предполагается наличие определенного языка, на котором происходит мышление. Правое полушарие реализует синтетический способ обработки информации, соединяя разрозненные сведения в единый образ. Оно действует как аналоговый компьютер, охватывая всю ситуацию сразу и реализуя существенный параллелизм. Правое полушарие мыслит на «языке» образов, структур. Происходит это значительно быстрее описания ситуации словами (аналитическим способом). Например, знакомое лицо человек распознает за 100–120 мс, цифровой компьютер тратит на это минуты и даже десятки минут (при наличии существенных помех). Нейрон состоит из тела (сомы) и отходящих от него отростков – дендритов и аксона. Сома содержит обычные для большинства клеток внутриклеточные компоненты: ядро и клеточные части, реализующие обмен веществ и синтез белков. Дендриты представляют собой сравнительно короткие ветвистые входные (афферентные) отростки. Они воспринимают идущие к нейрону по многочисленным волокнам импульсы (возбуждения). Аксон – это длинный, тонкий, мало ветвящийся (имеет концевые разветвления) выходной (эфферентный)отросток нейрона. Он передает нервные импульсы от нейрона к другим нервным клеткам, мышцам, железам. Пучки аксонов образуют нервные волокна. Типичная структура нейрона представлена на рис. 10 (в верхней его части). Следует заметить, что некоторые составляющие структуры нейрона, изображенные на рис. 10, оставлены без пояснения, т.к. не имеют принципиального значения для дальнейшего изложения материала. О структуре синапса, представленной в нижней части рисунка, речь пойдет несколько позже. Выявлено широкое разнообразие нейронов, различающихся формой тела, разветвленностью дендритной части и длиной аксона. Различают нейроны трех классов: · сенсорные (чувствительные, афферентные, рецепторные) нейроны, получающие информацию из внешней среды или от рецепторных клеток (фоторецепторов, механорецепторов и т.д.); · связующие (вставочные, ассоциативные) нейроны (интернейроны), которые связывают один нейрон с другим; · моторные (эффекторные, эфферентные) нейроны, посылающие свои импульсы к исполнительным органам (мышцам, железам).
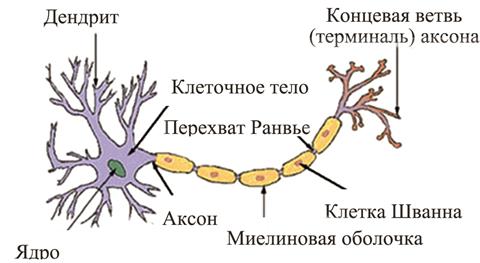
Рис. 10. Типичная структура нейрона и синапса
Головной мозг (высший отдел центральной нервной системы) почти целиком состоит из интернейронов. Различаясь строением, все нейроны одинаково проводят информацию. Они принимают входные сигналы, перерабатывают их и в виде нервных импульсов передают к эффекторным нервным окончаниям. Информация передается по аксонам в виде электрических импульсов (потенциалов действия) длительностью порядка 1 мс и амплитудой порядка 100 мВ. Сигнал, проходящий по аксону нервной клетки, формируется сложным образом. В состоянии покоя между внутренним содержимым нейрона и наружным раствором существует разность потенциалов (мембранный потенциал покоя) порядка 60—90 мВ, локализованная на поверхностной мембране. Внутренняя сторона мембраны заряжена отрицательно по отношению к наружной. Это обусловлено избирательной проницаемостью покоящейся мембраны для положительных ионов калия (К+), концентрация которых в протоплазме клетки в 50 раз выше, чем во внеклеточной жидкости. Диффузия К+ из клетки приводит к положительному заряду наружной стороны мембраны. При этом внутренняя сторона оказывается заряженной отрицательно. В то же время проницаемость покоящейся мембраны для положительных ионов натрия (Nа+) примерно в 100 раз ниже, чем для К+. И хотя Nа+ во внеклеточной жидкости в 10 раз больше, чем в протоплазме, их диффузия внутрь клетки мала, поэтому особо не влияет на потенциал покоя. Раздражители, действующие на клетку, деполяризуют мембрану, что приводит к увеличению ее проницаемости для Nа+. Поток натрия внутрь клетки еще сильнее деполяризует мембрану, увеличивая ее проницаемость и т.д. После некоторого критического (порогового) значения потенциала этот процесс приобретает взрывной характер – возникает потенциал действия. Его восходящая фаза на короткое время извращает мембранный потенциал (перебрасывает его знак на противоположный), после чего наступает нисходящая фаза, обусловленная падением проницаемости для Nа+ и ростом проницаемости для К+. Полное восстановление потенциала покоя наступает после окончания следовой деполяризации или следовой гиперполяризации мембраны, продолжительность которых существенно превосходит длительность пика. Соотношение концентраций Nа+ и К+ внутри и вне клетки поддерживается специальным механизмом – «натриево-калиевым насосом», выталкивающим ионы натрия из клетки и нагнетающим ионы калия внутрь клетки. Дендритная область и тело нейрона действуют совместно как интегрирующая система, накапливающая некоторую последовательность входных сигналов. В результате создается предпусковое состояние в области у основания аксона, называемой триггерной (пусковой) областью. Это наиболее возбудимый участок поверхностной мембраны нейрона, служащий местом возникновения нервного импульса. При достижении мембранным потенциалом в какой-то точке триггерной области порогового значения возникает импульс, распространяющийся по аксону без затухания. Нервный импульс – это нелинейная реакция нейрона на стимул, поддерживаемая за счет энергетических ресурсов аксона. Это движущаяся зона электрической деполяризации мембраны, которая распространяется по аксону со скоростью до 120 м/с благодаря токам действия, возникающим между возбужденным (деполяризованным) и соседним с ним покоящимся участками мембраны. Другими словами, нервный импульс распространяется по аксону путем регенерации потенциала действия. После прохождения импульса следует период абсолютной рефрактерности, в течение которого нейрон не может быть вновь возбужден ни при каком воздействии входных сигналов (эквивалентно заданию бесконечно большого порога). Суммарная длительность нервного импульса и периода абсолютной рефрактерности составляет около 1,5 мс (соответствует предельной частоте активизации нейрона порядка 666 Гц). Затем следует период относительной рефрактерности (несколько миллисекунд), в течение которого порог постепенно снижается до нормального уровня. Помимо рассмотренного механизма, в дендритах и теле клетки имеет место и другое взаимодействие полей, создаваемых потенциалами поступающих импульсов и распространяющихся по окружающей поверхности мембраны с некоторым затуханием. В зависимости от частоты поступающих импульсов и состояния воспринимающей мембраны в данный момент времени может происходить повышение ее чувствительности или прогрессирующее понижение. Таким образом, различные участки поверхностной мембраны оказываются специализированными каждый особым образом. Они по-разному реагируют на приходящие импульсы, а некоторые из них могут действовать как источники внутренней генерации сигналов и приводить к спонтанной активности нейрона. Единственный путь общения (взаимодействия) нейронов друг с другом – синапсы. Синапс (от греческого synapsis – соединение, связь) – это специализированный функциональный контакт между возбудимыми клетками, служащий для передачи и преобразования сигналов (нижняя часть рис. 10). Синапс включает пресинаптическую часть (синаптическое окончание, терминаль), синаптическую щель (порядка 20 нм), разделяющую контактирующие клетки, и постсинаптическую часть, т.е. участок клетки, к которому прилежит синаптическое окончание. Межнейронные синапсы могут быть образованы окончаниями аксонов одних нейронов и телом, дендритами или аксонами других нейронов. Соответственно различают аксо-соматические, аксо-дендритные и аксо-аксонные синапсы. Ввиду преобладания поверхности дендритов аксо-дендритные синапсы наиболее многочисленны. Механизм передачи через синапс может иметь химическую, физическую или смешанную природу. Наиболее распространены синапсы, реализующие связи между нейронами с помощью химических передатчиков – нейромедиаторов. Большинство нейромедиаторов – аминокислоты и их производные. Молекулы нейромедиатора выделяются из окончания аксона в синаптическую щель. Это происходит при достижении потенциалом действия окончания аксона. Высвободившиеся молекулы нейромедиатора связываются с рецепторами в постсинаптической мембране, изменяя ее проницаемость. При этом имеет место возбуждающий эффект, если мембрана деполяризуется, и тормозной, если мембранный потенциал стабилизируется на уровне потенциала покоя, либо происходит гиперполяризация мембраны. Для каждого нейрона характерен свой специфический тип медиатора. От тела большинства нейронов отходит много дендритов, с рецепторной мембраной которых контактирует множество окончаний аксонов других нейронов. Дендриты рецепторных нейронов способны трансформировать энергию внешнего раздражения в локальную импульсную активность. На мембране дендритов центральных нейронов происходит пространственно-временная суммация возбуждающих и тормозных постсинаптических потенциалов. В результате формируются нервные импульсы. Каждый синапс в отдельности лишь незначительно влияет на активность нейрона. Для установки интенсивности формирования нервного импульса нейрон должен непрерывно интегрировать до 1000 синаптических входов. Информация о записанных в памяти образах содержится в свойствах межнейронных синаптических контактов. Распознавание образа можно рассматривать как параллельный нейронный динамический процесс в сети от начального состояния, соответствующего входному образу, к конечному стабильному состоянию, отвечающему идеальному записанному образу. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
