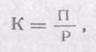|
|
ПОНЯТИЕ ОБ ЭКОСИСТЕМАХ. УЧЕНИЕ О БИОГЕОЦЕНОЗАХ
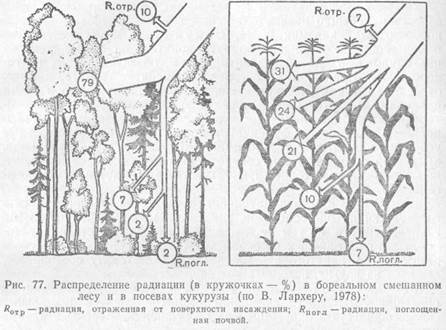
Сообщества организмов связаны с неорганической средой теснейшими материально-энергетическими связями. Растения могут существовать только за счет постоянного поступления в них углекислого газа, воды, кислорода, минеральных солей. Гетеротрофы живут за счет автотрофов, но нуждаются в поступлении таких неорганических соединений, как кислород и вода. В любом конкретном местообитании запасов неорганических соединений, необходимых для поддержания жизнедеятельности населяющих его организмов, хватило бы ненадолго, если бы эти запасы не возобновлялись. Возврат биогенных элементов в среду происходит как в течение жизни организмов (в результате дыхания, экскреции, дефекации), так и после их смерти, в результате разложения трупов и растительных остатков. Таким образом сообщество образует с неорганической средой определенную систему, в которой поток атомов, вызываемый жизнедеятельностью организмов, имеет тенденцию замыкаться в круговорот. Понятие об экосистемах. Любую совокупность организмов и неорганических компонентов, в которой может осуществляться кру-юворот веществ, называют экосистемой. Термин был предложен в 1935 г. английским экологом А. Тенсли, который подчеркивал, что при таком подходе неорганические и органические факторы выступают как равноправные компоненты и мы не можем отделить организмы от конкретной окружающей их среды. А. Тенсли рассматривал экосистемы как основные единицы природы на поверхности Земли, хотя они и не имеют определенного объема и могут охватывать пространство любой протяженности. Для поддержания круговорота веществ в системе необходимо наличие запаса неорганических молекул в усвояемой форме и трех функционально различных экологических групп организмов: продуцентов, консументов и редуцентов. Продуцентами выступают автотрофпые организмы, способные строить свои тела за счет неорганических соединений. Консументы — это гетеротрофные организмы, потребляющие органическое вещество продуцентов или других консументов и трансформирующие его в новые формы. Редуценты живут за счет мертвого органического вещества, переводя его вновь в неорганические соединения. Классификация эта относительная, так как и консументы, и сами продуценты выступают частично в роли редуцентов, в течение жизни выделяя в окружающую среду минеральные продукты обмена веществ. В принципе круговорот атомов может поддерживаться в системе и без промежуточного звена — консументов, за счет деятельности двух других групп. Однако такие экосистемы встречаются скорее как исключения, например на тех участках, где функционируют сообщества, сформированные только из микроорганизмов. Роль консументов выполняют в природе в основном животные, их деятельность по поддержанию и ускорению циклической миграции атомов в экосистемах сложна и многообразна. Масштабы экосистемы в природе чрезвычайно различны. Неодинакова также степень замкнутости поддерживаемых в них круговоротов вещества, т. е. многократность вовлечения одних и тех же атомов в циклы. В качестве отдельных экосистем можно рассматривать, например, и подушку лишайников на стволе дерева, и разрушающийся пень с его населением, и небольшой временный водоем, луг, лес, степь, пустыню, весь океан и, наконец, всю поверхность Земли, занятую жизнью. В подушке лишайников мы найдем все необходимые компоненты экосистемы. Продуценты — симбиотические водоросли, осуществляющие фотосинтез. В качестве консументов выступают некоторые мелкие членистоногие, питающиеся живыми тканями лишайника, а также грибные гифы, по существу паразитирующие на клетках водорослей. И гифы грибов, и большинство микроскопических животных, обитающих в лишайниковых подушках (клещи, коллемболы, нематоды, коловратки, простейшие), выступают и в роли редуцентов. Грибные гифы живут не только за счет живых, но и за счет погибших клеток водорослей, а мелкие животные-сапрофаги перерабатывают отмершие слоевища, в разрушении которых им помогают многочисленные микроорганизмы. Степень замкнутости круговорота в такой системе очень невелика: значительная часть продуктов распада выносится за пределы лишайника — вымывается дождевыми водами, осыпается вниз со ствола. Кроме того, часть животных мигрирует в другие местообитания. Тем не менее часть атомов успевает пройти несколько циклов, включаясь в тела живых организмов и освобождаясь из них, прежде чем покинет данную экосистему. В некоторых типах экосистем вынос вещества за их пределы настолько велик, что их стабильность поддерживается в основном за счет притока такого же количества вещества извне, тогда как внутренний круговорот малоэффективен. Таковы проточные водоемы, реки, ручьи, участки на крутых склонах гор. Другие экосистемы имеют значительно более полный круговорот веществ и относительно автономны (леса, луга, степи на плакорных участках, озера и т. п.). Однако ни одна, даже самая крупная, экосистема Земли не имеет полностью замкнутого круговорота. Материки интенсивно обмениваются веществом с океанами, причем большую роль в этих процессах играет атмосфера, и вся наша планета часть материи получает из космического пространства, а часть отдает в космос. В соответствии с иерархией сообществ жизнь на Земле проявляется и в иерархичности соответствующих экосистем. Экосистемная организация жизни является одним из необходимых условий ее существования. Запасы биогенных элементов, из которых строят тела живые организмы, на Земле в целом и на каждом конкретном участке на ее поверхности небезграничны. Лишь система круговоротов могла придать этим запасам свойство бесконечности, необходимое для продолжения жизни. Поддерживать и осуществлять круговорот могут только функционально различные группы организмов. Таким образом, функционально-экологическое разнообразие живых существ и организация потока извлекаемых из окружающей среды веществ в циклы — древнейшее свойство жизни. Учение о биогеоценозах. Параллельно с развитием концепции экосистем успешно развивается учение о биогеоценозах, автором которого был академик В. Н. Сукачев (1942). «Экосистема» и «биогеоценоз» — близкие по сути понятия, но если первое из них приложимо для обозначения систем, обеспечивающих круговорот любого ранга, то «биогеоценоз» — понятие территориальное, относимое к таким участкам суши, которые заняты определенными единицами растительного покрова — фитоценозами. Наука о биогеоценозах — биогеоценология — выросла из геоботаники и направлена на изучение функционирования экосистем в конкретных условиях ландшафта в зависимости от свойств почвы, рельефа, характера окружения биогеоценоза и составляющих его первичных компонентов — горной породы, животных, растений, микроорганизмов. Биогеоценология рассматривает поверхность Земли как сеть соседствующих биогеоценозов, связанных между собой через миграцию веществ, но тем не менее, хотя и в разной степени, автономных и специфичных по своим круговоротам. Конкретные свойства участка, занятого биогеоценозом, придают ему своеобразие, выделяя из других, исходных по типу. Обе концепции — экосистем и биогеоценозов — дополняют и обогащают друг друга, позволяя рассматривать функциональные связи сообществ и окружающей их неорганической среды в разных аспектах и с разных точек зрения. ПОТОК ЭНЕРГИИ
Поддержание, жизнедеятельности организмов и круговорот вещества в экосистемах возможны только за счет постоянного притока энергии. В конечном итоге вся жизнь на Земле существует засчет энергии солнечного излучения, которая переводится фотосинтезирующими организмами в химические связи органических соединений. Гетеротрофы получают энергию с пищей. Все живые существа являются объектами питания других, т. е. связаны между собой, энергетическими отношениями. Пищевые связи в сообществах — это механизмы передачи энергии от одного организма к другому. В каждом сообществе трофические связи переплетены в сложную сеть. Организмы любого вида являются потенциальной пищей многих других видов. Врагами тлей, например, служат личинки и жуки божьих коровок, личинки мух-сирфид, пауки, насекомоядные птицы и многие другие. За счет дубов в широколиственных лесах могут жить несколько сотен форм различных членистоногих, фитонематод, паразитических грибков и т.п. Хищники обычно легко переключаются с одного вида жертв на другой, а многие, кроме животной пищи, способны потреблять в некотором количестве и растительную. Таким образом, трофические сети в биоценозах очень сложные и создается впечатление, что энергия, поступившая в них, может долго мигрировать от одного организма к другому.
На самом деле путь каждой конкретной порции энергии, накопленной зелеными растениями, короток. Она может передаваться не более чем через 4—6 звеньев ряда, состоящего из последовательно питающихся друг другом организмов. Такие ряды, в которых можно проследить пути расходования изначальной дозы энергии, называют цепями питания. Место каждого звена в цепи питания называют трофическим уровнем. Первый трофический уровень—это всегда продуценты, создатели органической массы; растительноядные консументы относятся ко второму трофическому уровню; плотоядные, живущие за счет растительноядных форм,— к третьему; потребляющие других плотоядных — соответственно к четвертому и т. д. Таким образом, различают консументов первого, второго и третьего порядков, занимающих разные уровни в цепях питания. Естественно, что основную роль при этом играет пищевая специализация консументов. Виды с широким спектром питания могут включаться в пищевые цепи на разных трофических уровнях. Так, например, человек, в рацион которого входит как растительная пища, так и мясо травоядных и плотоядных животных, выступает в разных пищевых цепях в качестве консумента первого, второго и третьего порядков. Виды, специализированные на растительной пище, например тли, зайцеобразные, копытные, всегда являются вторым звеном в цепях питания. Энергетический баланс консументов складывается следующим образом. Поглощенная пища обычно усваивается не полностью.
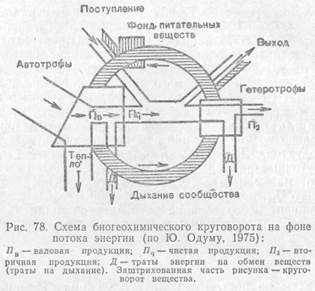
Неусвоенная часть вновь возвращается во внешнюю среду (в виде экскрементов) и в дальнейшем может быть вовлечена в другие цепи питания. Процент усвояемости зависит от состава пищи и набора пищеварительных ферментов организма. У животных усвояемость пищевых материалов варьирует от 12—20% (некоторые сапрофаги) до 75% и более (плотоядные виды). Ассимилированная организмом пища вместе с запасом в ней энергии расходуется двояким образом. Большая часть энергии используется на поддержание рабочих процессов в клетках, а продукты расщепления подлежат удалению из организма в составе экскрементов (мочи, пота, выделений различных желез) и_углекислого газа, образующегося при дыхании. Энергетические затраты на поддержание всех метаболических процессов условно называют тратой на дыхание, так как общие их масштабы можно оценить, учитывая выделение СО2 организмом. Меньшая часть усвоенной пищи трансформируется в ткани самого организма, т. е. идет на рост или откладывание запасных питательных веществ, увеличение массы тела. Эти отношения сокращенно можно выразить формулой: Р = П + Д + Н, где Ρ — рацион консумента, т. е. количество пищи, съедаемой им за определенный период времени, Π — продукция, т. е. траты на рост, Д — траты на дыхание, т. е. поддержание обмена веществ за тот же период, и Н — энергия неусвоенной пищи, выделенной в виде экскрементов. Передача энергии в химических реакциях в организме происходит согласно второму закону термодинамики, с потерей части ее в виде тепла. Особенно велики эти потери при работе мышечных клеток животных, КПД которых очень низок. В конечном итоге вся энергия, использованная на метаболизм, переходит в тепловую и рассеивается в окружающем пространстве. Траты на дыхание во много раз больше энергетических затрат на увеличение массы самого организма. Конкретные соотношения зависят от стадии развития и физиологического состояния особей. У молодых траты на рост могут достигать значительных величин, тогда как взрослые используют энергию пищи почти исключительно на поддержание обмена веществ и созревание половых продуктов. Интенсивность питания снижается с возрастом. Так, ежесуточный рацион карпов массой от 5 до 15 г составляет почти 1/4 от массы их тела, у более крупных особей — от 150 до 450 г—всего 1/10, а у рыб массой 500—800 г — 1/16. Коэффициент использования потребленной пищи на рост (К) рассчитывают как отношение этих двух величин:
где Π — траты на рост, Ρ — количество пищи, съеденной за тот же период. Двупарноногие многоножки кивсяки в период роста, который продолжается до трех лет, тратят на рост от 6 до 25% съеденной пищи при усвояемости в среднем 30%. В последующем их масса стабилизируется. Кивсяки живут до 12 лет. В умеренном поясе они активны 4—5 месяцев в году. Особь, масса которой во взрослом состоянии 0,5 г, за свою жизнь потребляет 250—300 г опада (80—90 г абсолютно сухой массы). Так как кивсяки многократно линяют, часть усвоенной энергии идет на восстановление покровов. Таким образом, отношение съеденного в течение жизни корма к массе взрослого животного составляет 500—600: 1. У такого гетеротермного животного, как малый суслик, который активен всего 2—2,5 месяца в году, это соотношение всего около 150: 1. Средний рацион суслика 30 г сухой массы растений (или в среднем 100 г сырой) при массе зверька 200 г и продолжительности жизни 4 года. Постоянно активным в течение года рыжим полевкам нужно гораздо больше энергии для поддержания жизнедеятельности. Взрослые зверьки массой 20 г съедают в среднем до 4 г сухого корма в день. При продолжительности жизни примерно в 24 месяца затрата кормов на жизнь одной особи составляет примерно 30 кг в сырой массе, что приблизительно в 1500 раз больше массы взрослого животного. Таким образом, основная часть потребляемой с пищей энергии идет у животных на поддержание их жизнедеятельности и лишь сравнительно небольшая — па построение тела, рост и размножение. Иными словами, большая часть энергии при переходе из одного звена пищевой цепи в другое теряется, так как к следующему потребителю может поступить лишь та энергия, которая заключается в массе поедаемого организма. По грубым подсчетам, эти потери составляют около 90% при каждом акте передачи энергии через трофическую цепь. Следовательно, если калорийность растительного организма 1000 Дж, при полном поедании его травоядным животным в теле последнего останется из этой порции всего 100, в теле хищника — лишь 10 Дж, а если этот хищник будет съеден другим, то на его долю придется только 1 Дж, т. е. 0,1%. Таким образом, запас энергии, накопленный зелеными растениями, в цепях питания стремительно иссякает. Поэтому пищевая цепь включает обычно всего 4—5 звеньев. Потерянная в цепях питания энергия может быть восполнена только поступлением новых ее порций. Поэтому в экосистемах не может быть круговорота энергии, аналогичного круговороту веществ. Экосистема функционирует только за счет направленного потока энергии, постоянного поступления ее извне в виде солнечного излучения или готовых запасов органического вещества. Трофические цепи, которые начинаются с фотосинтезирующих организмов, называют цепями выедания (или пастбищными, или цепями потребления), а цепи, которые начинаются с отмерших остатков растений, трупов и экскрементов животных,— детритными цепями разложения. Таким образом, поток энергии, входящий в экосистему, разбивается далее как бы на два основные русла, поступая к консументам через живые ткани растений или запасы мертвого органического вещества, источником которого также является фотосинтез. В разных типах экосистем мощность потоков энергии через цепи выедания и разложения различна: в водных сообществах большая часть энергии, фиксированной одноклеточными водорослями, поступает к питающимся фитопланктоном животным и далее—к хищникам и значительно меньшая включается в цепи разложения. В большинстве экосистем суши противоположное соотношение: в лесах, например, более 90% ежегодного прироста растительной массы поступает через опад в детритные цепи. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|