
|
|
Размножение организмов и геохимическая энергия живого вещества§ 25. Растекание размножением в биосфере зеленого живого вещества является одним из характернейших и важнейших проявлений механизма земной коры. Оно обще всем живым веществам, лишенным хлорофилла или им обладающим, оно — характернейшее и важнейшее выявление в биосфере всей жизни, коренное отличие живого от мертвого, форма охвата энергией жизни всего пространства биосферы. Оно выражается нам в окружающей природе во всюдности жизни, в захвате ею, если этому не препятствуют непреодолимые препятствия, всякого свободного пространства биосферы. Область жизни — вся поверхность планеты. Если какая-нибудь часть ее оказалась безжизненной, в короткий или медленный срок она неизбежно будет захвачена живыми организмами. Мера геологического времени в истории планеты — небольшой промежуток, и мы видим, как в это время вырабатываются организмы, приспособленные к жизни в условиях, которые раньше делали ее невозможной. Область жизни, по-видимому, расширяется в геологическом времени (§ 119, 122), и, во всяком случае, несомненно, что она всегда охватывает или стремится охватить до конца все доступное ей пространство — на протяжении, вероятно, всей геологической истории. Ясно, что это стремление является отличительной чертой живого вещества, а не проявлением чуждой ему силы, как, например, при растекании песчаной кучи или ледника под влиянием силы тяготения. Растекание жизни — движение, выражающееся во всюдности жизни — есть проявление ее внутренней энергии, производимой ею химической работы. Оно подобно растеканию газа, которое не есть следствие тяготения, но есть проявление отдельных движений частиц, совокупность которых представляет газ. Так и растекание по поверхности планеты живого вещества есть проявление его энергии, неизбежного движения, занятия нового места в биосфере новыми, созданными размножением организмами. Оно есть проявление прежде всего автономной энергии жизни в биосфере. Эта энергия проявляется в работе, производимой жизнью, в переносе химических элементов и в создании из них новых тел. Я буду называть ее геохимической энергией жизни в биосфере. § 26. Это движение живых организмов путем размножения, совершающееся с удивительной и неизменной математической правильностью, идет в биосфере непрерывно и является характернейшей и важнейшей по своим эффектам чертой ее механизма. Оно идет на земной поверхности — на суше, оно проникает все водоемы, в том числе гидросферу, оно видно на каждом шагу в тропосфере; в форме паразитов оно охватывает все другие живые существа, имеет место внутри самих живых веществ. Неуклонно и неизменно оно длится без перерыва и без замедления мириады лет, все время совершая огромную геохимическую работу, являясь формой проникновения энергии солнечного луча в нашу планету и ее распределения по земной поверхности. Мы должны видеть в нем не только перенос материальных тел, но и передачу энергии. В связи с этим и перенос материальныхтел размножением есть процесс особенный. Это не есть простое механическое передвижение тел по земной поверхности, независимых, не связанных с той средой, в которой они двигаются. Среда, в которой они движутся, не только обусловливает своим сопротивлением трение, как это имеет место в движении тел под влиянием тяготения. В этом движении связь со средой глубже — оно может идти только под влиянием газового обмена движущихся тел и той среды, в которой происходит движение. Оно тем быстрее, чем газовый обмен сильнее, оно замирает, когда газовый обмен не может иметь места. Газовый обмен — это дыхание организмов; оно, как мы увидим, глубоко меняет, направляет размножение. В движении размножения мы видим проявление его геохимического значения, выражение его как части механизма биосферы, подобно тому как само это движение является отражением солнечного луча. Проявлением энергии того же луча является и само дыхание — газовый обмен жизни с окружающей средой. § 27. Хотя это движение идет кругом нас непрерывно, мы его не замечаем, так как мы нашим взором охватываем только общий его результат — ту красоту, разнообразие форм и красок, движений и соотношений, которые нам дает живая природа. Мы видим лишь поля и леса с их растительной и животной жизнью, полные жизни водоемы и моря, пронизанную ею только кажущуюся мертвой почву. Мы видим статический результат, динамическое равновесие этих движений; редко когда можем наблюдать их как таковые. Остановимся на нескольких примерах, в которых вскроется это созидающее живую природу, нам не видное, но для живой природы основное своеобразное движение. Временами на небольших относительно пространствах мы видим прекращение жизни высшей растительности. Лесной пожар, степные палы, взрыхленные, распаханные, запущенные поля, вновь образовавшиеся острова, застывшие потоки лавы, покрытые вулканическим пеплом пространства суши, освободившиеся от ледников или водных бассейнов ее пространства, новые почвы, образованные на безжизненных скалах лишайниками и мхами, — эти и другие формы бесконечных проявлений жизни на нашей планете на некоторое время образуют лишенные трав и деревьев пятна на зеленом покрове суши. Они образуют их на короткое время. Жизнь быстро входит в свои права. И зеленые травы, а затем и древесная растительность занимают утерянные или новые места. Отчасти они занимают их проникновением извне, приносом семян подвижными организмами или еще больше — ветром, отчасти возникают от всюду в почве находящихся их запасов, лежащих в ней в латентном состоянии и сохраняющихся в этой форме иногда, по крайней мере, столетия. Но это проникновение семян извне есть необходимое условие заселения, но не оно его производит. Заселение осуществляется благодаря размножению организмов, зависит от специфичной для их размножения геохимической энергии; оно идет годами, пока не будет восстановлено нарушенное равновесие. Как мы увидим, это находится в полном соответствии со скоростью передачи в биосфере жизни, передачи геохимической энергии этих живых веществ — высших зеленых растений, В этом случае, следя внимательно за заселением пустых пространств, человек может видеть движение растекания жизни, о котором я говорю, реально ощущать ее давление; вдумываясь в него, он может созерцать движение на нашей планете солнечной энергии, превращенной в земную — химическую. Он его ощущает и в тех случаях, когда ему приходится защищать от чуждого заселения нужные ему поля или пустые пространства, тратить на преодоление давления жизни свою энергию. Он видит его и тогда, когда всматривается в окружающую его природу, в глухую, молчаливую, беспощадную борьбу за существование, которую ведут кругом него зеленые растения. Он видит и ощущает действительно, как надвигается лес на степь или как лишайниковая тундра в своем движении его глушит. § 28. Членистоногие насекомые, клещи, пауки составляют главную массу живого животного вещества суши. В тропических и подтропических странах среди них преобладающую роль играют Orthoptera, муравьи, термиты. Размножение их идет своеобразным путем. Хотя геохимическая энергия, ему отвечающая (§ 37), и того же порядка, как для высших зеленых растений, она все же несколько меньше. В государствах термитов дает потомство, производит непосредственно размножение, один организм из десятков тысяч, иногда сотен тысяч бесполых неделимых. Это царица-мать. Она кладет яички непрерывно весь свой век — иногда десять и больше лет. Количество яичек, новых особей, которое она может дать, исчисляется биллионами. В год она дает сотни тысяч. Указываются случаи, когда она дает 60 яичек в минуту, т. е. 86 400 штук в сутки, так же правильно, как в часовых механизмах отбиваются секунды, суточное количество которых равно тем же 86 400. Размножение идет роями. Часть потомства с новой царицей-маткой улетает и захватывает новое пространство вне ареала, необходимого для жизни первого, исходного государства. Всюду действует с математической точностью инстинкт: и в сохранении яичек, немедленно уносимых термитами-рабочими, и в отлете роев, и в замене — в случае неожиданных случайностей — старой матки новой. Всюду действует с той же точностью и число. Все подвержено мере, числовой закономерности: число яичек, число годовых роев, в них неделимых, число населения государств, размеры и вес организмов, темп размножения и вызванного им переноса геохимической энергии термитов по земной поверхности. Мы можем точно выразить числом напряжение движения термитов по земной поверхности благодаря их размножению, зная годовое количество роев, число в них особей, их размеры, количество яичек, отлагаемых в год царицей; мы можем обнять числом отвечающее этому движению его отражение в окружающей среде, его давление. Давление это очень велико. Человек, живущий в области их обитания, знает это по той работе, которую он должен производить, чтобы защищать от них продукты своего существования, своего питания. Если бы не было препятствий во внешней среде, главным образом в окружающей термитов жизни, в немногие годы они могли бы захватить и покрыть своими государствами всю поверхность биосферы — 5,10065 • 108 км2. § 29. Среди организмов бактерии занимают особое место. Это организованные тела мельчайших известных размеров: линейные размеры их измеряются 10 - 4 см и даже 10 -5 см. В то же самое время это организмы с наибольшей силой размножения. Они размножаются дроблением. Каждая клетка многократно удваивается в сутки. Наиболее быстро размножающаяся бактерия производит эту работу 63 - 64 раза в сутки, в среднем каждые 22 - 23 минуты, с такой же правильностью, как откладывает яички самка термитов или обращается около Солнца планета, на которой эта бактерия живет. Бактерии живут в жидкой или полужидкой среде. Главные их массы наблюдаются в гидросфере; значительные количества сосредоточены в почве, проникают в другие организмы. Если бы не было препятствий во внешней среде, они могли бы создать с непостижимой для нас быстротой невероятные количества сложнейших химических соединений, являющихся вместилищем огромной химической энергии. Этой огромной энергии отвечает огромная быстрота их размножения. Таким путем в течение полутора и менее суток бактерии могли бы покрыть тонким однослойным покровом поверхность земного шара, которую в результате размножения зеленые травы или насекомые могли бы преодолеть в течение ряда лет, в отдельных случаях сотен дней. В морской среде находятся бактерии почти шаровой формы, объем которых, по М. Фишеру, достигает 1 Процесс деления неизбежно происходит таким темпом, если бактерия живет в условиях жизни, этому благоприятных, прежде всего, если температура среды это дозволяет. Если температура падает, быстрота чередования поколений уменьшается, и это изменение может быть выражено в точной числовой формуле. Все время бактерия дышит, т. е. находится в тесной связи с растворенными в воде газами. Ясно, что количество бактерий путем размножения никогда не может достигнуть в 1 см3 той величины, которая определяет в нем количество газовых молекул, т. е. 2,706 · 1019 (число Лошмидта). Газовых молекул в 1 см3, заполненном водой, будет во много раз меньше. Мы видим здесь предел размножению ставящийся явлениями дыхания, свойствами газообразного состояния материи. §30. Пример бактерий позволяет выразить движение, наблюдаемое в биосфере благодаря размножению, в другой форме, чем мы это делали до сих пор. Представим себе период в истории Земли, который гипотетически неправильно, как увидим, допускают геологи, время, когда океан покрывал не три четверти земной поверхности, а всю планету. Э. Зюсс относил это «вселенское море» (по-гречески «Панталасса») в археозойскую эру. Бактерии в это время, несомненно, его населяли. Их следы известны в слоях древнейшего палеозоя. Характер минералов археозойских слоев и, особенно характер их ассоциаций с неменьшей несомненностью доказывают нахождение бактерий во всем археозое — в самых древнейших, доступных геологическому изучению пластах нашей планеты. Если бы в этом «вселенском море» температура была благоприятна для их жизни и если бы в нем не было препятствий их размножению, шаровая бактерия, в 10-12 см3 объемом, в 1, 47 суток — меньше чем в полутора суток — образовала бы в этом море сплошную пленку в 5,10065 · 108 км2. Пленки бактерий, образующиеся благодаря их размножению, занимают хотя и меньше, но все же весьма значительные площади в биосфере. В 90-х годах прошлого столетия проф. М. А. Егунов указывал на существование тонкой пленки серных бактерий на всей площади Черного моря. Эта пленка равнялась бы в таком случае поверхности Черного моря, т. е. 411 540 км2, и лежала бы на границе кислородной поверхности, т. е. на глубине 200 м. Однако исследования проф. Б. Л. Исаченко, участника экспедиции Н. М. Книповича (1926), не подтверждают этих данных. Это же явление в менее грандиозных размерах, но отчетливо выражено в динамических равновесиях живых организмов, например на границе между пресной и соленой водой и Мертвом озере Кильдинских островов, которое на всей своей поверхности всегда покрыто сплошной пленкой пурпурных бактерий (К. Дерюгин, 1926). Другие микроскопические, но все же более крупные организмы постоянно дают примеры подобных явлений. От времени до времени пленка, образованная этими организмами океанического планктона, покрывает пространство в тысячи квадратных километров. Подобные пленки образуются довольно быстро. Во всех этих случаях можно изобразить геохимическую энергию этих процессов таким же образом, а именно в виде передвижения этой энергии на земной поверхности, причем скорость растения пропорциональна скорости размножения вида, в нашем случае бактерии Фишера. При своем максимальном развитии, когда данный вид заселит всю земную поверхность (5,10065 • 108 км2), эта энергия за определенное время, разное для каждого вида, пройдет также максимальное расстояние, равное земному экватору, т. е. 40 075 721 м. Бактерия Фишера, размером в 10-12 см3, при образовании пленки во «вселенском море» Э. Зюсса развила бы энергию, скорость продвижения которой по земному диаметру была бы равна около 33 100 см/с. Это явление может быть выражено в иной форме. Скорость V, равная 33 100 см/с, может быть рассматриваема как скорость передачи жизни, геохимической энергии, вокруг земного шара; она равна средней скорости вращения вокруг него бактерии путем размножения. В 1,47 суток бактерия размножением обтекает земной шар, совершает вокруг него во «вселенском море» полный оборот... Скорость передачи жизни, по наибольшему расстоянию, ей доступному, величина V, будет той характерной для каждого однородного живого вещества постоянной, которой мы будем пользоваться для выражения геохимической энергии жизни. §31. В основе этой величины, всегда отличной для всякого вида или расы, сказываются, с одной стороны, характер механизма размножения, а с другой стороны, те пределы возможному размножению, которые кладутся размерами и свойствами планеты. Скорость передачи жизни не есть простое выражение свойств автономных организмов или их совокупностей — живых веществ; она выражает их размножение в соответствии с биосферой как планетное явление. В ее выражение неизбежно входят элементы планеты — величины ее поверхности и ее экватора. Мы имеем здесь аналогию с некоторыми другими свойствами организма, например с его весом. Вес организма на Земле и того же организма на другой планете будет иной, хотя организм может при этом не измениться. Точно так же и скорость передачи его жизни, например, на Земле или Юпитере, площадь и экватор которого иные, чем Земли, будет иная, хотя бы сам организм оставался при этом неизменным. Этот специфически земной характер скорости передачи жизни вызывается тем ограничением, которое свойства и характер Земли как планеты, биосферы как космического явления вносят в проявление заложенного в организмах как в автономных созданиях механизма размножения. § 32. Область явлений размножения мало обращала на себя внимание биологов. Но в ней — отчасти незаметно для самих натуралистов — установилось несколько эмпирических обобщений, которые отчасти кажутся нам сами по себе понятными, так мы с ними свыклись. Среди этих обобщений необходимо отметить следующие: 1) Размножение всех организмов выражается геометрическими прогрессиями. Можно выразить это в единообразной формуле:
где п — число дней с начала размножения;
Характерным для каждого живого вещества является Процесс мыслится бесконечным, как бесконечной является прогрессия. 2) Эта бесконечность возможности проявления размножения организма сказывается в подчинении этого проявления в биосфере, т. е. растекания живого вещества, правилу инерции. Может считаться эмпирически установленным, что процесс размножения задерживается в своем проявлении только внешними силами; он замирает при низкой температуре, прекращается или ослабляется при недостатке пищи или дыхания, при отсутствии места для обитания вновь создаваемых организмов. Уже в 1858 г. Ч. Дарвин и А. Уоллес высказали эту мысль в форме, которая была давно ясна натуралистам, вдумывавшимся в эти явления, например К. Линнею, Ж. Бюффону, А. Гумбольдту, К. Эренбергу, К. М. Бэру: если не будет внешних препятствий, всякий организм в разное, определенное для него время может размножением покрыть весь земной шар, произвести по объему потомство, равное массе океана или земной коры. 3) Темп размножения, сказывающийся в таком эффекте, отличен для каждого организма и находится в тесной зависимости от размеров организма. Мелкие организмы, т. е. организмы в то же время и более легкие, размножаются гораздо быстрее, чем большие организмы, т. е. организмы в то же время большего веса. § 33. В этих трех эмпирических положениях явления размножения организмов выражены вне времени и пространства или, вернее, в геометрических и механических бесформенных однородных времени и пространстве. В действительности жизнь в той форме, в какой мы ее изучаем, есть чисто земное — планетное — явление, не отделимое от биосферы, созданное и приспособившееся к ее условиям. Перенесенная в отвлеченное время и отвлеченное пространство математики жизнь является фикцией, созданием нашего разума, отличным от реального явления. Если мы хотим иметь точные, научные представления в наши положения о ее свойствах, мы должны внести поправки в отвлеченные понятия времени и пространства; эти поправки могут в корне, как мы видим в данном случае, изменить наши выводы, в которых свойства земных времени и пространства не были предусмотрены. § 34. На Земле организмы живут в ограниченном пространстве, одинаковом по размерам для всех них. Они живут в пространстве определенного строения в газообразной или проникнутой газами жидкой среде. И хотя время нам представляется безграничным, но время какого-нибудь процесса в ограниченном пространстве, каким является размножение организмов, не может являться безграничным. Оно тоже будет иметь предел, различный для каждого организма в зависимости от характера его процесса размножения. Неизбежным следствием этого положения является ограничение всех величин, определяющих явления размножения организмов в биосфере. Должны существовать наибольшие числа неделимых, которые могут дать разные живые вещества. Эти числа — Nmax — должны быть конечны и характерны для каждого вида и расы. Скорости передачи жизни должны заключаться в точных и определенных пределах, которые не могут быть никогда превзойдены. Наконец, величины Эти пределы устанавливаются двумя проявлениями планеты: 1) ее размерами и 2) физическим заполнением пространства, в котором течет жизнь, жидкостями и газами — прежде всего свойствами газов и характером газового обмена. § 35. Остановимся на ограничении, вносимом размерами планеты. Влияние этих размеров мы видим на каждом шагу. Небольшие водоемы очень часто покрыты сплошь на своей поверхности плавающей на них зеленой растительностью. В наших широтах это очень часто зеленые ряски — разные виды Lemna. Поверхность воды представляет сплошной зеленый их покров без промежутков. Растеньица тесно сдвинуты, их зеленые пластинки заходят друг на друга; процесс размножения действует, но он замедлен внешним препятствием — прежде всего отсутствием места. Он проявляется только тогда, когда в зеленом покрове вследствие внешних разнообразных причин — гибели рясок или их уноса — образуются пустые промежутки водной поверхности. Они немедленно замешаются размножением. Очевидно, количество неделимых ряски, могущих поместиться на данной площади, определенно и находится в зависимости от размеров и условий их существования. Когда оно достигнуто, процесс размножения останавливается, задерживается внешним неодолимым препятствием. В каждом пруду создается своеобразное динамическое равновесие, аналогичное тому, какое в нем наблюдается при испарении воды с его поверхности. Упругость паров и упругость жизни механически аналогичны. Другой всем известный пример представляет в картине природы жизнь зеленой водоросли — разных видов Protococcus, обладающей гораздо большей геохимической энергией, чем ряска. Она покрывает в благоприятных условиях сплошь без промежутков (§ 50) стволы деревьев. Дальше ей идти некуда; ее процесс размножения задержан; он возобновляется вновь, как только открывается возможность помещения новых неделимых Protococcus. Количество неделимых этой водоросли, могущих поместиться на площади дерева, строго определенно и не может быть превзойдено. § 36. Эти соображения могут быть целиком перенесены на всю живую природу и на область, доступную ее обитанию— на поверхность нашей планеты. Наибольшее могущее существовать проявление силы размножения живого вещества определено размерами планеты и выражается в количестве неделимых, которые могут разместиться на площади, равной 5,10065 • 108 км2. Это количество есть функция густоты скопления организмов, возможной для их жизни. Эта густота очень различна; для ряски или одноклеточной водоросли протококка она определяется только их размерами; другие организмы требуют гораздо большей площади (или объема) для жизни. Слон в Индии требует до 30 км2, овца на горных пастбищах Шотландии — около 10 5 м2, средний улей пчел — не менее 10 - 15 км2 (одна пчела — не менее 2 • 10 5 км2, т. е. 200 м2) среднего красного леса Украины, от 3000 до 15 000 неделимых планктона хорошо развиваются в 1000 см3 морской воды, 25 - 30 см2 достаточны для обычных злаков, несколько (иногда десятков) квадратных метров — для неделимых обычного нашего леса. Очевидно, скорость передачи жизни зависит от возможной густоты хорошо живущей, не страдающей в своих проявлениях совокупности неделимых, от плотности живого вещества. Не буду здесь останавливаться на очень еще мало изученной этой важной константе жизни в биосфере. Ясно, что наибольшая плотность сплошного покрова (типа ряски или протококка) или сплошного заполнения 1 см3 мельчайшей бактерией (§ 29) даст нам, если принять ее возможной для всех организмов, наибольшее допустимое для данного вида количество его неделимых в биосфере. Для получения этого числа необходимо принять плотность равной квадрату максимального изменения организма, т. е. его длины и ширины (коэффициент k1)[4]. § 37. Ограничение размножения размерами планеты, неизбежная остановка процесса уже этим путем, помимо более глубокого влияния, оказываемого, как увидим, зеленой средой (§ 123), придает этому процессу очень своеобразные и важные черты. Прежде всего, очевидно, есть предельное, одинаковое для всех организмов, наибольшее расстояние, по какому может распространяться передача жизни. Оно равно земному экватору, т.е. 40 075 721 м. Во-вторых, для всякого вида или расы есть максимальное количество неделимых, которое не может быть никогда превзойдено. Это максимальное число получается при полном заполнении данным видом земной поверхности при максимальной возможной густоте его обитания. Это число, которое я буду дальше обозначать Nmax и называть стационарным числом однородного живого вещества, имеет большое значение для оценки геохимического влияния жизни. Оно отвечает максимально возможному проявлению энергии данного однородного живого вещества в биосфере, максимальной его геохимической работе; скорость его достижения, разная для каждого организма, выражается скоростью V — передачи жизни. Эта скорость — V — связана со стационарным числом следующей формулой:
§ 38.Мы видим это явление в окружающей нас природе. Оно давно было замечено старыми натуралистами и ярко подчеркнуто около сорока лет назад точным наблюдателем живой природы. К.Земпером (1888). Земпер отметил, что в небольших водоемах при всех равных условиях размножение организмов уменьшается по мере увеличения в них количества неделимых. Стационарное число не достигается или достижение его замедляется по мере приближения к нему количества создаваемых организмов; существуют какая-то причина, может быть не всегда внешняя (§43),регулирующая процесс. Опыты Пирля и его сотрудников над мухой Drosophila и над курами (1911 - 1912) подтверждают это обобщение Земпера в другой среде. § 39. Скорость передачи жизни может давать нам ясное понятие о геохимической энергии жизни разных организмов. Она колеблется в больших пределах и находится в тесной зависимости от размера организма. Для самых мелких организмов, для бактерий, она, как мы видели, близка к скорости звука, т. е. к величине, равной 33 100 см/с. Для самых крупных, для крупных млекопитающих, она равна долям сантиметра — для индийского слона, например, V= 0, 09 см/с. Это крайние пределы. Между ними помещаются скорости передачи жизни для всех других организмов. Они находятся в явной зависимости от размеров организма и в более простых случаях (например, для организмов, форма которых приближается к шару) связь размеров организма с его скоростью v может быть уже сейчас математически выражена. Но существование определенной математической зависимости всегда и везде в этой области несомненно и отвечает старинному прочному эмпирическому обобщению (§32). § 40. Скорость передачи жизни дает ясное понятие об энергии жизни в биосфере, о ее в ней работе, но оно недостаточно для ее определения. Для этого мы должны принять во внимание массу того организма, энергия растекания совокупностей которого в биосфере определяется скоростью V. Выражение скорость растекания геохимической энергии которого равна V, дает нам выражение кинетической геохимической энергии живого вещества. Взятое по отношению к определенным площади или объему биосферы, оно может дать нам выражение той химической работы, которая в геохимических процессах этой площади или объема может быть произведена данным видом или расой организмов. Уже давно мы имеем подходы к определению этим путем по отношению к определенной площади биосферы— гектару— части геохимической энергии живого вещества. Это делается при определении урожаев — количества с данной площади полезных человеку организмов или их продуктов. В более полной форме оно выражается в количестве органического вещества, которое может быть создано размножением и ростом организмов на гектаре. Хотя эти данные очень неполны и не охвачены теорией в достаточной степени, они привели уже к важным эмпирическим обобщениям. Несомненно, количество создаваемого на гектаре органического вещества ограничено и теснейшим образом связано Во-вторых, все более выясняется, что в случаях максимальных урожаев количества органического вещества с гектара почвы и гектара океана суть числа одного порядка, приближающиеся к одной и той же величине. Гектар суши охватывает ничтожный слой, не превышающий метров, гектар океана отвечает слою воды, захваченной жизнью, измеряемому километрами. Тождественность создаваемой в них энергии жизни, очевидно, указывает на освещение сверху как на ее источник. Мы увидим, что, вероятно, это связано с характерным свойством почвы суши, скопляющей в себе концентрацию организмов, обладающих огромной геохимической энергией (§ 155). Благодаря этой концентрации энергии живого вещества тонкий слой почвы по своему геохимическому эффекту может сравниваться с огромной толщей моря, где центры жизни разжижены инертной массой воды. §41. Кинетическая геохимическая энергия организма
Чрезвычайно характерно, что эта величина для Protozoa есть величина постоянная. Для них всех выражение A1, принимает форму:
где а — коэффициент, близкий к единице[6]. Из этой формулы ясно, что кинетическая геохимическая энергия Protozoa определяется скоростью V, связанной с весом, размерами организма и темпом размножения в которой числовые, постоянные для всех видов организмов коэффициенты связаны с размерами планеты (с площадью ее поверхности и длиной ее экватора, причем величины выражены в сантиметрах и секундах)[8]. Из формулы скорости очевидно, что одни размеры планеты не могут объяснить действительно существующего предела для V и Наибольшая нам известная величина V равна 33 100 см/с, а наибольшая величина Могут ли они идти дальше (что, как это видно из приведенных формул, возможно и при постоянстве кинетической энергии на гектар) или есть в биосфере условие, этому препятствующее? Такое условие есть, и им является газовый обмен организмов, неизбежный и необходимый для их существования и, в частности, для их размножения. § 42. Организм не существует без газового обмена, без дыхания. Чем размножение идет быстрее, тем дыхание становится интенсивнее. По степени газового обмена мы всегда можем судить об интенсивности жизни. В масштабе биосферы нам необходимо, конечно, иметь в виду не дыхание отдельного организма, а общий результат дыхания, необходимо учесть газовый обмен — дыхание всех живых организмов, обнять его как часть механизма биосферы. В этом отношении давно имеются эмпирические обобщения, очень мало до сих пор обращавшие на себя внимание и не учтенные нашей научной мыслью. Одно из них указывает, что газы биосферы те же, которые создаются при газовом обмене живых организмов. В биосфере существуют только они одни: О2, N2, СО2, Н2О, Н2, СН4, NH3... Это не может быть случайностью. Затем весь свободный кислород биосферы создается на земной поверхности только благодаря газовому обмену зеленых организмов. Этот свободный кислород есть главный источник свободной химической энергии биосферы. И, наконец, в-третьих, количество этого свободного кислорода в биосфере, равное 1,5 • 1021 г, есть число того же порядка, как и количество существующего и с ним неразрывно связанного живого вещества, исчисляемого в 102О - 1021 г. Оба эти исчисления получены вполне независимо друг от друга. Эта тесная связь газов Земли с жизнью указывает с несомненностью, что газовый обмен организмов — и на первом месте их дыхание — должен иметь первостепенное значение в газовом режиме биосферы, т. е. являться планетным явлением. § 43. Этот газовый обмен — дыхание — определяет весь темп размножения; он ставит пределы для величин V и Они не могут перейти через пределы, нарушающие свойства газов. Я уже указывал (§ 29), что количество организмов, могущих существовать в одном кубическом сантиметре среды, должно быть меньше количества в нем газовых молекул, т. е. меньше 2, 706·1019 (число Лошмидта). Если величина v будет больше 33 100 см/с, то для организмов, размеры которых меньше размеров бактерий (т. е. размеров меньшего порядка, чем п •10-5), количество неделимых благодаря их размножению превысит 1019 в 1 см3. Очевидно, при неизбежном существовании обмена между газовыми молекулами и организмами количество поглощающих и выделяющих газовые молекулы организмов, соизмеримых с молекулами по своим размерам, должно было бы расти по мере уменьшения размеров организмов с все большей, в конце концов, невероятной быстротой. По нашим современным представлениям, таким образом, приходим к физическому абсурду. Если ограничение количества неделимых в кубическом сантиметре определяет наименьшие размеры организма и этим путем ставит максимальный предел для 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
 3, т. е. 10-12 см3. В 1 см3 может заключаться 1012 неделимых, и при быстроте их размножения — в сутки около 63 делений каждой клетки — кубический сантиметр может быть ими заполнен в течение нескольких — 11 - 13 — часов, если в него попадет одна такая бактерия. В действительности бактерии живут не изолированно, а образуют колонии и при благоприятных условиях они заполняют 1 см3 еще быстрее.
3, т. е. 10-12 см3. В 1 см3 может заключаться 1012 неделимых, и при быстроте их размножения — в сутки около 63 делений каждой клетки — кубический сантиметр может быть ими заполнен в течение нескольких — 11 - 13 — часов, если в него попадет одна такая бактерия. В действительности бактерии живут не изолированно, а образуют колонии и при благоприятных условиях они заполняют 1 см3 еще быстрее.
 — показатель прогрессии, который для одноклеточных организмов, размножающихся делением, соответствует числу поколений в сутки;
— показатель прогрессии, который для одноклеточных организмов, размножающихся делением, соответствует числу поколений в сутки;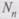 — число неделимых, существующих благодаря размножению через п дней.
— число неделимых, существующих благодаря размножению через п дней. В этой формуле никаких пределов, никаких ограничений ни для п, ни для
В этой формуле никаких пределов, никаких ограничений ни для п, ни для  ни для N нe заключается.
ни для N нe заключается. геометрических прогрессий размножения тоже имеют определенные пределы.
геометрических прогрессий размножения тоже имеют определенные пределы.
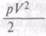 , где р — средний вес организма[5],
, где р — средний вес организма[5], с той солнечной лучистой энергией, которую захватывает зеленое растение. Геохимическая энергия, собранная этим путем — размножением организмов, — есть измененная солнечная энергия.
с той солнечной лучистой энергией, которую захватывает зеленое растение. Геохимическая энергия, собранная этим путем — размножением организмов, — есть измененная солнечная энергия. - количество организмов на гектар при достижении ими стационарного числа (§ 37), а к — коэффициент плотности жизни (8 36):
- количество организмов на гектар при достижении ими стационарного числа (§ 37), а к — коэффициент плотности жизни (8 36):

 Отнесенное к
Отнесенное к  , V выражается
, V выражается  следующей простой формулой[7]:
следующей простой формулой[7]:
 — около 63 - 64.
— около 63 - 64.
 и V, то неизбежное постоянное соотношение между количеством неделимых и газовых молекул в данном объеме — явления дыхания — играет еще большую и постоянно проявляющуюся роль в явлениях размножения.
и V, то неизбежное постоянное соотношение между количеством неделимых и газовых молекул в данном объеме — явления дыхания — играет еще большую и постоянно проявляющуюся роль в явлениях размножения.