|
|
Эволюционная теория изо дня в день подтверждается научно-исследовательской практикойПрактикующих биологов убеждают в реальности эволюции не только (и даже не столько) факты, изложенные в предыдущих разделах (не каждый биолог держит их постоянно в голове), сколько повседневная научно-исследовательская практика, которая, как известно, критерий истины. Эволюционная теория — весьма эффективное практическое средство, позволяющее предсказывать новые, еще не открытые факты, целенаправленно искать их и находить. В предыдущих разделах мы уже упоминали о нескольких предсказаниях такого рода, которые направили работу исследователей в определенную сторону (т.е. стали исследовательской программой) и привели к успеху, то есть к открытию новых фактов, прежде неизвестных, но предсказанных теорией. К числу самых известных примеров относится предсказанная Дарвином длительная история жизни на Земле в период, предшествовавший «кембрийскому взрыву« — и триумфальный успех палеонтологии докембрия в XX веке; предсказанное и подтвержденное основателями молекулярной генетики единство генетического кода всех живых организмов (см. раздел «Молекулярно-генетические и биохимические доказательства«), многочисленные предсказанные и найденные ископаемые переходные формы (например, тиктаалика искали совершенно целенаправленно в отложениях строго определенного возраста, там, где была «дырка» между уже известными переходными формами от рыб к четвероногим), см. раздел «Палеонтологические доказательства«. Но убежденность биологов основана не столько на этих хрестоматийных и широко разрекламированных триумфах эволюционной теории, сколько на бесчисленном множестве открытий меньшего масштаба, но зато повседневных и повсеместных, которые были выполнены по алгоритму «предсказал — проверил — подтвердил». Рассмотрим пару примеров, взятых наугад из статей, опубликованных в течение последнего месяца (эти строки написаны в феврале 2010 г.)
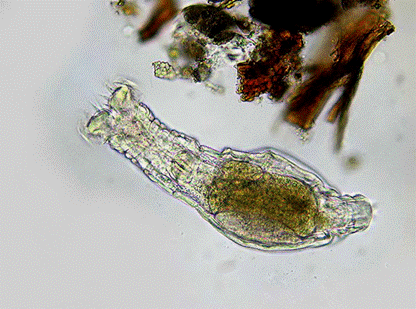
1. Как коловратки спасаются от паразитов.Эволюционная теория позволила дать убедительный ответ на очень непростой вопрос о том, почему в природе так широко распространено половое размножение (ведь гораздо проще почковаться или размножаться партеногенетически, как некоторые организмы и делают). Суть этого ответа в том, что половое размножение резко повышает эффективность естественного отбора, ускоряя отбраковку вредных и накопление полезных мутаций. Этот эффект полового размножения был предсказан теоретически, а затем подтвержден экспериментально. История о том, как эволюционная теория позволила понять, зачем нужно половое размножение, тоже очень подходит нам для иллюстрации основной идеи данного раздела (подробности см. в заметке «Опыты на червях доказали, что самцы — вещь полезная«), но мы сейчас хотим рассказать о другом, более частном исследовании. Способность полового размножения ускорять адаптивную (приспособительную) эволюцию особенно важна для борьбы с паразитами, которые ведут со своими жертвами нескончаемую «эволюционную гонку вооружений» (см.: Современные паразиты опаснее прошлых и будущих). Бделлоидные коловратки — единственная большая группа животных, в которой утрата полового размножения не привела к быстрому вымиранию. Все остальные группы, отказавшиеся от полового размножения, представляют собой маленькие концевые веточки (или небольшие пучки веточек) на эволюционном дереве. Это означает, что такие группы обычно вымирают, не успев дать начало большим таксонам (семействам, отрядам или классам). Теория предсказывает, что важной причиной вымирания бесполых популяций являются паразиты, обгоняющие их в эволюционной гонке и доводящие до полного истребления. Заметьте, вот еще одна маленькая деталь, показывающая силу эволюционной теории. Мы берем одну эволюционную модель (объяснение, зачем нужно половое размножение), выводим из нее проверяемое следствие (группы животных, отказавшиеся от полового размножения, должны в большинстве случаев быстро вымирать), и затем проверяем это предположение при помощи другой эволюционной модели (в данном случае это многочисленные независимые методы построения эволюционных деревьев). Если обе наши модели правильные, мы должны увидеть, что большинство бесполых групп животных представляют собой отдельные концевые веточки на эволюционном дереве или маленькие концевые пучки веточек. Именно это мы и видим. Тем самым добавляется еще одна капелька в давно уже переполненную чашу нашей уверенности в реальности эволюции. Бделлоидные коловратки, однако, обходятся без полового размножения уже как минимум 40 миллионов лет. За это время они успели стать довольно разнообразной группой, которой биологи придают ранг класса и которая включает более 450 современных видов. Следовательно, если теория верна, у коловраток должно быть какое-то «тайное оружие» против паразитов, позволяющее им процветать невзирая на отсутствие полового размножения. Биологи Кристофер Уилсон и Пол Шерман из Корнеллского университета (США) предположили, что бделлоидные коловратки спасаются от паразитов благодаря трем своим уникальным свойствам. Во-первых, бделлоидные коловратки умеют переносить полное высыхание, причем жизнеспособность в высушенном состоянии у них сохраняется до 9 лет. Во-вторых, в сухом виде они очень легко переносятся ветром (именно поэтому многие виды бделлоидных коловраток имеют глобальное распространение). В-третьих, они моментально «отмокают» и начинают расти и размножаться, стоит им попасть в любое влажное место, будь то эфемерная маленькая лужа или пучок сырого мха. Главными врагами бделлоидных коловраток являются паразитические грибы рода Rotiferophthora. Это специализированные паразиты, вызывающие смертельное заболевание у бделлоидных коловраток и ни у кого больше. Гриб убивает коловратку, прорастает наружу через ее покровы и рассеивает споры, которые поедаются другими коловратками и заражают их. Заболевание характеризуется 100-процентной смертностью — это как раз то, чего и следует ожидать в случае проигранной эволюционной гонки.
Слева — здоровая бделлоидная коловратка, справа — то, что от нее остается после заражения грибом Rotiferophthora
Исследователи решили проверить, могут ли коловратки избавиться от паразитов при помощи высыхания и последующего перенесения ветром. Коловраток рассаживали по одной в чашки Петри и давали им спокойно жить и размножаться в течение 9 дней. За это время каждая коловратка успевала дать начало маленькой популяции из нескольких десятков особей. После этого во все чашки, кроме контрольных, добавляли споры смертоносного гриба Rotiferophthora angustispora. Гриб начинал бурно размножаться и истреблял всех коловраток в чашке за 12–15 дней. Затем ученые попробовали высушивать зараженную популяцию коловраток через три дня после заражения и оставлять в сухом виде на неделю. После этого в чашку снова наливали воды. Поначалу казалось, что всё идет нормально: коловратки оживали и выглядели здоровыми, а вода была незаразной для других коловраток, что свидетельствует об отсутствии живых грибных спор. Но уже через 48 часов из тел мертвых коловраток начинали показываться гифы уцелевших грибов, они рассеивали споры, и через 2–3 недели популяция коловраток погибала. Оставив зараженных коловраток в высушенном виде не на одну, а на две недели, ученые получили точно такой же результат: после добавления воды оживали не только коловратки, но и грибы, и всё кончалось плачевно. Однако результаты получились совсем другими, когда популяции высушивали на более долгий срок — 3, 4 или 5 недель. В этом случае большинство «размоченных» популяций оставались свободными от грибной инфекции вплоть до конца эксперимента (в течение 20 недель). После трехнедельного высушивания грибы «ожили» только в 40% подопытных популяций, после четырехнедельного — в 15%, пятинедельного — в 9,5%. Таким образом, коловратки выдерживают высыхание лучше, чем их паразиты. Ясно, что эта способность должна помогать им бороться с грибной инфекцией в природе. Помогает ли им в этом также и распространение при помощи ветра? Ученые соорудили камеру, в которой дул легкий ветерок, и поставили туда чашку с зараженной и только что высушенной популяцией коловраток. В чашке были также естественные субстраты: мох и ил. На расстоянии 30–40 см были установлены 24 пустые чашки. За неделю в эти чашки нанесло ветром в общей сложности 5 мг сухого материала. Чашки залили водой, и в 17 из них вскоре обнаружились ожившие коловратки. В семи из этих семнадцати популяций вскоре появился гриб-паразит, который расправился с коловратками, как обычно, в течение 2–3 недель. Однако остальные 10 популяций оказались свободными от гриба и чувствовали себя нормально до окончания эксперимента. Таким образом, без ветра коловраткам требовалось пролежать в сухом виде не менее трех недель, чтобы избавиться от инфекции. Переносимые ветром, коловратки освобождались от гриба гораздо быстрее. Следовательно, ветер действительно может помогать коловраткам спасаться от паразита. Видимо, это связано с тем, что коловратки лучше приспособлены к распространению при помощи ветра, чем гриб. Здоровые коловратки, высыхая, превращаются в компактные (менее 300 мкм) и очень легкие овальные тельца, которые называют «бочонками» (tuns). «Бочонки» исключительно хорошо переносятся ветром. Гриб же приспособлен в основном к распространению через воду. В контрольном эксперименте, когда зараженную популяцию не высушивали и расселяли капельками воды, все дочерние популяции оказались зараженными и вскоре погибли. Очевидно, что в природе коловраткам и их паразитам часто приходится сталкиваться и с более долгими засухами, и с переносом на гораздо большие расстояния, чем в проведенных экспериментах. Полученные результаты показали, что коловратки ухитряются выжить без полового размножения благодаря тому, что хорошо научились играть с паразитами в прятки. Таким образом, предсказанное эволюционной теорией «тайное оружие» коловраток было успешно найдено благодаря целенаправленному поиску.Источник:Christopher G. Wilson, Paul W. Sherman. Anciently Asexual Bdelloid Rotifers Escape Lethal Fungal Parasites by Drying Up and Blowing Away // Science. 2010. V. 327. P. 574–576.
2. Взаимопомощь среди сперматозоидов.Полиандрия — спаривание самки со многими самцами — приводит к острой конкуренции между сперматозоидами за право оплодотворить яйцеклетку. Эту конкуренцию можно рассматривать с двух точек зрения: отдельных сперматозоидов (с одинарным набором хромосом) и целых самцов (с двойным набором). Эволюционная гонка вооружений, порождаемая «спермовыми войнами», ведет к выработке специфических адаптаций на обоих уровнях. Как правило, самцов тех видов, для которых характерны спермовые войны, легко опознать по крупным семенникам. Чем больше они производят спермы, тем выше их шансы оставить потомство (типичный пример — шимпанзе; у человека, напротив, маленькие семенники — это свидетельствует о том, что в эволюции гоминид спермовые войны не имели большого значения). Сперматозоиды, со своей стороны, тоже вырабатывают специфические приспособления для победы в отчаянной гонке, в которой генам-победителям достается шанс на бессмертие. Одним из таких приспособлений является способность сперматозоидов объединяться в группы, склеиваясь «головка к головке» или «головка к хвосту». Смысл в том, что такие группы быстрее движутся в половых путях самки и обгоняют конкурентов-одиночек. В кооперации сперматозоидов присутствует элемент альтруизма, ведь из всей группы только один сперматозоид может передать свои гены следующим поколениям, а все остальные жертвуют собой, чтобы ему помочь. Но они, скорее всего, не знают заранее, кто именно окажется счастливчиком, а у сперматозоидов-одиночек шансы на успех всё равно ниже, чем у любого члена группы. Однако при полиандрии выгодно помогать не всякому сперматозоиду, а только родственнику, то есть сперматозоиду того же самца. Сперматозоиды одного самца имеют в среднем по 50% одинаковых генов, поэтому, согласно правилу Гамильтона (см.: Эволюция кооперации и альтруизма), каждому сперматозоиду выгодно пожертвовать частью собственных шансов на успех ради родственников, если их суммарный выигрыш от этого более чем вдвое превысит его проигрыш. Совсем другое дело, если речь идет о чужих сперматозоидах: помогать им во вред себе совершенно невыгодно. Таким образом, эволюционная теория предсказывает, что у видов, для которых характерна полиандрия, отбор может поддержать способность сперматозоидов отличать родню от чужаков и кооперироваться только со «своими». Для проверки этого предсказания ученые из Гарвардского университета (США) изучили поведение сперматозоидов у двух близкородственных видов американских хомячков рода Peromyscus, которых в Америке называют «оленьими мышами» (deer mice) (см. о них в заметке: Эволюция защитной окраски у мышей изучена на молекулярном уровне). Для одного из видов (P. maniculatus) характерна полиандрия, причем между спариваниями одной самки с двумя разными самцами проходит порой меньше минуты. Самки второго вида (P. polionotus) более целомудренны и предпочитают устойчивые семейные связи. В соответствии с этим и семенники у самцов P. maniculatus втрое крупнее, чем у P. polionotus. Для начала исследователи убедились, что у обоих видов сперматозоиды склонны объединяться в группы. Эти группы, как показали эксперименты in vitro (в пробирке), плавают в среднем со скоростью 127 микрометров в секунду и уверенно обгоняют одиночек, средняя скорость которых составляет лишь 110 мкм/с. После этого были проведены эксперименты на избирательность объединений. Сперматозоиды разных особей красили двумя разными флюоресцентными красителями (красным и зеленым), а затем следили за их поведением в искусственной среде. В качестве контроля в этих опытах использовались сперматозоиды одного самца, половина из которых была выкрашена в красный, половина — в зеленый цвет. Подсчитывался процент одноцветных сперматозоидов в каждой группировке. Оказалось, что если смешать сперматозоиды двух видов, то они образуют преимущественно конспецифичные группировки, то есть объединяются в соответствии с видовой принадлежностью. Значит, способность к различению «своих» и «чужих» у сперматозоидов имеется. Во втором эксперименте смешали сперматозоиды двух разных самцов P. maniculatus. Они тоже образовали одноцветные группы по родственному принципу. Но когда экспериментаторы смешали сперматозоиды двух самцов моногамного вида P. polionotus, для которых спермовые войны не актуальны, сперматозоиды стали образовывать смешанные красно-зеленые группировки. Таким образом, эксперименты полностью подтвердили теоретические ожидания: у вида с полиандрией сперматозоиды разных самцов различают друг друга и объединяются только со «своими», а у моногамного вида сперматозоиды не имеют такой избирательности. Как и в предыдущем примере, неизвестный ранее факт — умение сперматозоидов группироваться по родственному принципу — был обнаружен в результате целенаправленного поиска. Эволюционная теория подсказала исследователям, что искать и где, и привела их к успеху. Источник: Heidi S. Fisher, Hopi E. Hoekstra. Competition drives cooperation among closely related sperm of deer mice // Nature. Advance online publication 20 January 2010. Постоянно сталкиваясь с подобными ситуациями, биологи раз за разом убеждаются в действенности эволюционных моделей. Стоит ли удивляться, что среди биологов сегодня практически не осталось антиэволюционистов?
Cui prodest? «Кому выгодно?» — один из полезнейших принципов сыскного дела. Он приложим и к биологии. Эволюционная теория предсказывает, что адаптации (приспособления) организмов должны быть выгодны в первую очередь генам (аллелям), отвечающим за эти признаки. «Выгода» в эволюции означает «репродуктивный успех», то есть успешное размножение. Если признак, определяемый данным аллелем, способствует более успешному распространению данного аллеля в популяции (по сравнению с другими, конкурирующими аллелями, определяющими альтернативные варианты данного признака), то аллель будет распространяться (будет «поддержан отбором») и в конце концов может вытеснить конкурирующие аллели из генофонда популяции. Таким образом, в самом простом «базовом» случае эволюция работает строго в интересах «эгоистичного гена». Однако гены почти всегда действуют не поодиночке, а слаженными «коллективами». Каждый такой коллектив — геном отдельного организма — имеет общие интересы, которые состоят в том, чтобы несущий их организм оставил как можно больше жизнеспособного и плодовитого потомства. Поэтому эволюция обычно (почти всегда) работает также и в интересах «эгоистичной особи». Иногда складываются более сложные ситуации, когда «эгоистичному гену» выгодно пожертвовать одной или двумя своими копиями ради того, чтобы повысить репродуктивный успех других своих копий. В этом случае эволюция начинает работать в интересах другой особи или группы особей (подробнее см.: Эволюция кооперации и альтруизма). Таким образом, предсказания эволюционной теории относительно того, кому должны быть выгодны адаптации, напоминают три закона роботехники Айзека Азимова: 1) Адаптация должна быть выгодна генам, которые ее определяют (кодируют). 2) Адаптация должна быть выгодна особи, несущей эти гены в своем геноме, если это не противоречит первому закону. 3) Адаптация должна быть выгодна родным и близким данной особи, если это не противоречит первому закону (противоречие второму закону допускается). Это, конечно, крайне упрощенные и не совсем корректные формулировки, но в рамках популярного текста их можно рассматривать как допустимые. Из всего этого следуют довольно интересные проверяемые следствия, подтверждаемые многочисленными фактами. Например, теория предсказывает, что могут существовать «обнаглевшие» эгоистичные гены (или небольшие компании генов), действующие исключительно в своих корыстных интересах и в ущерб интересам того организма, в котором они находятся. Это может наблюдаться только в том случае, если ген имеет возможность размножаться независимо от размножения целого организма (иначе их эволюционные интересы будут полностью совпадать — как оно обычно и бывает — и тогда «эгоизм» гена не сможет проявиться). За примерами далеко ходить не надо — это, прежде всего, вирусы. Есть веские основания полагать, что по крайней мере некоторые группы вирусов — это «одичавшие» фрагменты геномов сложных организмов (см.: Данные сравнительной геномики проливают свет на происхождение ретровирусов). К этой же группе явлений относятся и плазмиды (см.: Plasmid). Как вирусы, так и плазмиды часто вступают в симбиоз со своими хозяевами, становятся безвредными или начинают приносить пользу, но только если это выгодно им самим, улучшает их шансы на успешное размножение. В принципе вирусу — при прочих равных — только лучше, если зараженный им организм будет жить долго и счастливо. Вся проблема в оговорке «при прочих равных». Если долгую жизнь хозяина можно купить только ценой снижения эффективности распространения вируса, то вирус на это не пойдет (то есть отбор такую мутацию не поддержит). Теория также предсказывает, что взаимопомощь и альтруизм должны гораздо чаще встречаться между близкими родственниками, чем между неродственными представителями одного и того же вида. Природа буквально переполнена живыми подтверждениями этого предсказания (многочисленные примеры подробно разобраны в уже неоднократно цитировавшемся обзоре «Эволюция кооперации и альтруизма«).
Почему среди перепончатокрылых так распространен альтруизм? Один из ярких примеров действенности эволюционных моделей — это объяснение, данное Уильямом Гамильтоном тому удивительному факту, что из всех животных почему-то только у перепончатокрылых насекомых (муравьев, пчел, ос, шмелей) много раз независимо возникала эусоциальность (eusociality) — общественный образ жизни, основанный на полном отказе большинства членов группы от размножения. Почему именно перепончатокрылые обладают повышенной склонностью к отказу от собственного размножения ради воспитания сестер? Разгадка кроется в особенностях наследования пола в этом отряде насекомых. У перепончатокрылых самки имеют двойной набор хромосом и развиваются из оплодотворенных яиц. Самцы гаплоидны (имеют одинарный набор хромосом) и развиваются из неоплодотворенных яиц. Из-за этого складывается парадоксальная ситуация: сестры оказываются более близкими родственницами, чем мать и дочь. У большинства животных степень родства между сестрами и между матерями и дочерьми одинакова (50% общих генов). У перепончатокрылых родные сестры имеют 75% общих генов, потому что каждая сестра получает от отца не случайно выбранную половину его хромосом, а весь геном полностью. Мать и дочь у перепончатокрылых имеют, как и у других животных, лишь 50% общих генов. Вот и получается, что для эффективной передачи своих генов следующим поколениям самкам перепончатокрылых, при прочих равных, выгоднее заставлять свою мать рожать новых сестер и помогать ей их выращивать, чем обзаводиться собственным потомством. Многие факты убедительно свидетельствуют о том, что альтруизм перепончатокрылых целиком и полностью основан на «генетическом эгоизме» (см.: Склонность к альтруизму сильнее у тех, кому нечего терять; Альтруизм общественных насекомых поддерживается полицейскими методами. Эволюционная теория утверждает, что «генетический альтруизм» невозможен в принципе, по определению. Иными словами, естественный отбор не может закрепить мутацию, снижающую эффективность распространения мутировавшего аллеля по сравнению с его конкурентами — аллелями того же гена, не имеющими этой мутации. Это невозможно даже в том случае, если мутация выгодна для популяции или вида в целом. Аллель, который попытается вести себя «альтруистично» по отношению к другим аллелям того же гена, будет автоматически «отбракован» естественным отбором. С другой стороны, те виды, в которых эгоистическая эволюция аллелей зайдет слишком далеко, с большой вероятностью просто вымрут. Поэтому в биоте постепенно накапливаются такие виды, у которых по тем или иным причинам интересы эгоистичных генов не слишком сильно расходятся с интересами вида как целого. Но отбор внутри популяции, отбор на уровне особей, работает гораздо быстрее и эффективнее, чем отбор на уровне видов. Любая адаптация должна сначала сформироваться на уровне популяции, и для этого соответствующие аллели должны «победить» своих конкурентов — альтернативные аллели тех же генов. Иначе адаптация просто не закрепится, не станет нормой для данного вида. И только уже закрепившиеся адаптации попадают в сферу внимания «межвидового отбора» (избирательного вымирания) — процесса гораздо более медленного, и потому обладающего гораздо меньшей «созидательной силой». Поэтому теория эволюции предсказывает, что: (1) в большинстве случаев адаптации должны быть полезны одновременно и для генов, которые их кодируют, и для особей, и для вида в целом, и даже, как правило, для всей экосистемы; (2) довольно часто могут встречаться ситуации, когда адаптации выгодны для генов, которые их кодируют, но вредны для вида и (или) для экосистемы; (3) обратные ситуации, когда адаптация полезна для вида, но вредна для распространения аллелей, «кодирующих» данную адаптацию, должны встречаться очень редко или не встречаться вовсе. А что «предсказывает» антиэволюционизм? Он, как обычно, не предсказывает вообще ничего до тех пор, пока не будут хоть как-то конкретизированы представления о намерениях «разумного дизайнера». Если исходить из предположения, что «дизайнер» имел в виду создание хорошей, устойчиво функционирующей биосферы, то он должен был бы проектировать в первую очередь адаптации, выгодные биосфере как целому, затем отдельным экосистемам, затем, возможно видам и популяциям. Совершенно непонятно, зачем бы ему создавать адаптации, выгодные «эгоистичному гену» или «эгоистичной особи» и вредные для популяции, вида или экосистемы. Таким образом, идея «благонамеренного разумного дизайнера» дает отчасти такие же, а отчасти иные предсказания, чем теория естественного отбора. Предсказание (1) в обоих случаях совпадает, здесь мы не можем различить научную и антиэволюционистскую модели по их следствиям. Однако в предсказаниях (2) и (3) появляются очевидные различия. Как обычно, факты явно на стороне научной модели. Рассмотрим несколько характерных примеров.
Почему соотношение полов почти всегда 1:1? У подавляющего большинства раздельнополых животных поддерживается соотношение полов, очень близкое к 1:1. Это вполне рационально для моногамных видов, у которых образуются устойчивые брачные пары и самцы участвуют в заботе о потомстве вместе с самками. Однако моногамия — весьма редкое явление в животном мире. Гораздо больше существует видов, у которых о потомстве заботятся только самки, а роль самца ограничивается спариванием. Например, самцы павлинов тратят кучу энергии на отращивание своих прекрасных хвостов, едят пищу, которая пригодилась бы самкам и птенцам, короче говоря, только зря топчут землю и занимают жизненное пространство. Животноводам прекрасно известно, что на большое стадо коров вполне достаточно одного быка, всех остальных самцов можно смело пускать на мясо. Никакого вырождения породы от этого не происходит — скорее наоборот. В природе у стадных копытных самцы могут быть полезны для коллективной обороны стада от хищников, но гораздо чаще самцы не делают и этого. Даже если рассмотреть виды, у которых от самцов есть какая-то польза, кроме производства спермы, все равно нет абсолютно никаких оснований полагать, что при любом образе жизни, способе питания, экологической обстановке и т.п. соотношение полов 1:1 является оптимальным для любого вида животных. Если бы соотношение полов определялось интересами вида, мы бы наверняка наблюдали разное соотношение полов у разных видов. Однако мы за редчайшими исключениями везде наблюдаем одно и то же соотношение 1:1 — у моногамных и полигамных видов, у заботящихся о потомстве и у тех, кто бросает потомство на произвол судьбы, у хищников и их жертв, у тех, кто организует коллективную оборону от хищников, и тех, кто этого не делает, и т.д. Это совершенно непохоже на разумный дизайн и необъяснимо с позиций пользы для вида или биосферы. Эволюционная теория, напротив, дает четкое и красивое объяснение этому феномену. Дело в том, что равное соотношение полов является в подавляющем большинстве случаев единственной эволюционно стабильной стратегией (Evolutionarily stable strategy). Это значит, что только при таком соотношении полов никакая мутация, меняющая эта соотношение, не окажется выгодной для ее носителя. Рассмотрим воображаемую ситуацию, когда у некого вида самки рожают не 50% детенышей мужского пола, а, скажем, только 20%. Соответственно, в популяции на каждого самца будет приходиться 4 самки. Допустим, что это соотношение является оптимальным с точки зрения «интересов вида». Но такая ситуация эволюционно нестабильна, и вот почему. В нашей воображаемой популяции хоть самцов и меньше, чем самок, каждый детеныш все равно имеет только одну мать и только одного отца. Допустим, популяция состоит из 1000 особей, из них 200 самцов и 800 самок, и каждый год в популяции рождается 1000 детенышей. Тогда получается, что каждая самка оставит в среднем 1000/800=1.25 детенышей, а каждый самец — 1000/200=5 детенышей. Самцы размножаются в 4 раза эффективнее! Иными словами, в такой популяции быть самцом в 4 раза выгоднее, чем самкой. Теперь посмотрим, что произойдет, если в популяции появится мутантный аллель, носительницы которого рожают больше детенышей мужского пола, чем остальные самки — например, не по одному сыну на каждые четыре дочери, а по одному сыну на две дочери. Очевидно, что самка, несущая этот аллель, оставит больше внуков, чем прочие самки, и, следовательно, мутантный аллель начнет распространяться в популяции. Вскоре он неизбежно вытеснит конкурирующий (старый, «немутантный») аллель, заставляющий самок рожать детей в соотношении 1:4. В итоге соотношение полов в популяции изменится: теперь самцов будет не вчетверо, а только вдвое меньше, чем самок. Нетрудно увидеть, что подобные мутации будут до тех пор менять соотношение полов, пока оно не станет равным 1:1. Только при таком соотношении никакая новая мутация, меняющая число рождаемых мальчиков и девочек, уже не станет полезной для самки и не будет поддержана отбором. Итак, в популяции установится соотношение полов 1:1, и никого не волнует, что вид в целом от этого только проиграет. Вид может в конце концов даже вымереть — что ж, тем хуже для него. «Межвидовой отбор» слишком медлителен и слаб, чтобы воспрепятствовать действию отбора на уровне аллелей в популяции. На самом деле виды не вымирают из-за неоптимального соотношения полов прежде всего потому, что это их общая проблема — они все в таком положении. Соотношение полов 1:1 — это почти неизбежный побочный эффект раздельнополости, следствие конкуренции между эгоистичными генами (особями), не имеющий отношения к благу вида или биосферы. Упомянутые выше «редкие исключения» из общего правила только подтверждают его. Например, у некоторых насекомых (бабочек, мух, жуков) доля самцов в популяции может быть резко снижена из-за деятельности весьма любопытной паразитической бактерии вольбахии. Вольбахия живет внутри клеток насекомого-хозяина и не может передаваться горизонтально, т.е. заражать других насекомых. Вместо этого она передается вертикально, то есть наследуется, причем только по материнской линии. Она проникает из материнского организма в яйцеклетки, и таким образом передается потомству зараженной самки. В сперматозоид бактерия проникнуть не может — он слишком мал, и поэтому бактерии, живущие в самце, обречены погибнуть вместе с ним. С точки зрения бактерии, хозяева-самцы — это тупик, западня. Поэтому «эгоистичные интересы» вольбахии состоят в том, чтобы в популяции хозяев стало побольше самок и поменьше самцов. Вольбахия «научилась» достигать этой цели, избирательно убивая эмбрионы мужского пола. В результате в некоторых зараженных популяциях насекомых на сотню самок может приходиться всего 1-2 самца. Как ни странно, такие популяции, судя по всему, чувствуют себя отлично и вовсе не собираются вымирать. Некоторые эксперты предполагают, что эти популяции на самом деле выигрывают от того, что вольбахия помогает им поддерживать оптимальное соотношение полов. Сами насекомые не могут этого добиться из-за конфликта интересов своих «эгоистичных генов», но бактерия приходит на помощь (см. наш обзор: Бактерия вольбахия — повелитель мух).
Другие примеры адаптаций, бесполезных либо вредных для вида, но полезных для эгоистичного гена (или эгоистичной особи). Почему деревья такие высокие? Они тянутся к свету, и высокий рост позволяет им не оказаться затененными другими деревьями. Если бы деревья могли договориться между собой, они бы, возможно, ввели закон: не расти выше стольких-то метров. И им не пришлось бы тратить непомерное количество ресурсов на производство древесины. Но деревья не могут договориться, и поэтому любая мутация, позволяющая дереву вырасти чуть выше других, поддерживается отбором (потому что дерево, приподнявшееся над лесным пологом, получит больше света, произведет больше семян и оставит больше потомства). В итоге новая мутация распространится, и все деревья станут чуть выше. Эта бессмысленная (с точки зрения «блага вида») гонка продолжается до тех пор, пока увеличение роста не наткнется на какую-либо эволюционную преграду, так что дальнейший рост будет уже не улучшать а ухудшать шансы дерева на успешное размножение (например, возрастет вероятность быть поваленным ураганом). Почему самцы так редко помогают самкам заботиться о потомстве? Если уж соотношение полов должно быть равным, и в популяции поэтому много «лишних» самцов, почему бы их не привлечь к делу? Зачем им без толку слоняться и только тратить на свое бесполезное для вида существование ценные пищевые ресурсы? Для эволюционной теории тут нет никакого парадокса: самцам часто бывает выгоднее не тратить силы и время на уход за детенышами, а заниматься поиском и соблазнением новых самок (а для этого, между прочим, полезно иметь много сил, выглядеть здоровым, хорошо питаться и поменьше работать). В первом случае самец только повышает шансы на выживание небольшого числа детенышей, во втором — у него появляется возможность произвести этих детенышей в десятки раз больше. Конечно, в каждой конкретной ситуации выгодность той или иной стратегии — заботливого отца или «мачо» — будет зависеть от конкретных обстоятельств (например, от того, насколько сильно отцовская забота может снизить смертность детенышей). Характерно, что у некоторых животных «эволюционно стабильным состоянием» оказывается одновременное существование двух типов самцов: заботливых отцов и бродячих бездельников-совратителей. Вторые, по сути, паразитируют на альтруизме первых (см.: Бойцы и проныры имеют равные шансы на успех в личной жизни;Владение земельной собственностью снижает плодовитость). К адаптациям, выгодным «эгоистичному гену» или «эгоистичной особи», но бесполезным или вредным для вида в целом, относится большинство адаптаций, связанных с «эволюционным конфликтом полов», и значительная часть адаптаций, связанных с «эволюционными гонками вооружений» (см. раздел «Этические и эстетические аргументы«). Несомненно вреден для вида (но, к сожалению, полезен для эгоистичных самцов) инфантицид — убийство самцами чужих детенышей, о чем тоже рассказано в разделе «Этические и эстетические аргументы«. Ведь при этом убиваются не наименее приспособленные, не больные и не ущербные, а нормальные детеныши, вся «вина» которых заключается только в том, что они не родные дети своему «отчиму». Выше мы рассмотрели пример, связанный со взаимопомощью сперматозоидов. При этом мы оставили в стороне один интересный вопрос: зачем у моногамного вида, у которого нет полиандрии и, соответственно, нет конкуренции между сперматозоидами разных самцов, сперматозоиды все равно объединяются в группы? По-видимому, потому, что сперматозоид-мутант, который не захочет ни с кем объединяться, просто-напросто проиграет гонку другим сперматозоидам того же самого самца. В данном случае налицо адаптация, не нужная ни виду в целом, ни даже отдельным самцам. Она выгодна только «эгоистичному сперматозоиду». Опять-таки, это совсем непохоже на «разумный дизайн», зато очень похоже на результат действия естественного отбора.
* * * Существуют и другие группы фактов, демонстрирующих явное превосходство эволюционной теории над идеей «разумного дизайна» как по предсказательной, так и по объяснительной силе. Возможно, в будущем мы добавим в эту публикацию разделы, посвященные побочным эффектам эволюции, компромиссам (trade-offs) и другим явлениям, явно выдающим почерк естественного отбора, а вовсе не «разумного дизайна». 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|