
|
|
Уравнение Ходжкина Хаксим.Впервые материальная модель распределения нервного импульса в аксоне была предложена в 1961 г. Ходжкином и Хаксимом. При построении математической модели распределения импульсов в активной среде необходимо учитывать изменение проводимости мембран для ионов калия, натрия и некоторых других видов ионов. Для этого в модели использовались проводимость мембран для различных ионов. Построение модели опиралось на два предложения: 1. Ионы калия проникают через мембрану при сдвиге потенциала до критического льшь тогда, когда вблизи калиевого канала присутствуют 4 активирующие этот процесс частицы.
h – вероятность подхода к каналу одной активной части. 2. Ионы натрия проникают через мембрану лишь в том случае, если в близи канала находится 3 активных частицы и одна блокирующая.
m – вероятность подхода к каналу одной активной частоты. n – вероятность подхода к каналу одной блокирующей частицы. На основании экспериментальных данных, а именно за счёт измерения токов через мембрану гигантского аксона при различных напряжениях, было установлено, что суммарный ток мембраны складывается из следующих токов:
Суммарный ток возникающий за счёт других ионов.
Т.о. полный ток
Т.о. ток через мембрану зависит от приложенного напряжения и от вероятности подхода активирующих и блокирующих частиц к ионным каналом. Для их определения была использована следующая система уравнений составленная из законов кинетики:
Нервное волокно можно представить в виде эквивалентной элементарной схемы с распределёнными параметрами в качестве активного сопротивления, увеличивающего амплитуду сигналов вводится генератором ионов K, Na и др. т.е. схема может быть представлена:
Уравнение распространения нервного импульса вдоль такой линии с некоторой константой скоростью описывается волновым уравнением:
Для распределения импульса, вдоль волокна который соответствует форме нервного волокна. Сила тока в волноводе имеет вид:
х – координата, вдоль которой, распространяется волна.
Объединяя полученное выражение, для силы тока Х и х получим уравнение для определения скорости:
В соответствии с уравнением теоретически получено значение скорости распространения, экспериментально получим значение
Синоптическая передача. Декрементный и не декрементный механизм передачи возбуждения характерен для распространения нервного импульса в пределах 1 клетки. Переходящее возбуждение от одной клетки на другую требует особого дополнительного механизма. Такой переход получил название синоптической передачи. Существует 2 вида синоптической: 1. Электрический. 2. Химический. Электрическая, СП возможна лишь при плотном контакте двух клеток, величина зазора между клетками не может превышать 10-15нм. В этом случае процесс возбуждения может передаваться за счёт перехода электромагнитных волн от одной мембраны на другую, т.е. происходит возбуждение ПД в мембране второй клетки за счёт потенциала первой клетки – это есть электрический синопс . Однако, электрический синопс встречается довольно редко, не более 1-3% от общего числа передач. В большинстве контактов расстояние между мембранами превышает 20 нм и электрический синапс оказывается невозможен даже при наличии щелевых контактов, что связано с быстрым затуханием волны в проводящей среде. В этом случае в передаче возбуждения участвующие химические посредники - медиатор. В первые гипотеза о химическом синопсе была сформулирована в 1924 году Самойловым А.Ф. в последствии многократно подтверждена экспериментами. Синоптическая передача состоит из трёх основных элементов:
1-е и 2-е принадлежат к 1-ой клетки, 3-е ко 2-ой клетки. Медленный поток медиаторов через постоянную структуру не возбуждает клетку. Движение медиатора резко ускоряется при деполяризирующем мембранном потенциале. При этом открывается Са каналы, что приводит к преступлению ионов Са+ в структуру из межклеточной среды. Избыток Са+ понижает заряд мембранных пузырьков, содержащих медиаторы и тем самым способствует перестройке присиноптической структуры, которая в результате пропускает большее количество медиаторов. Т.о. нервный импульс ускоряет экзоцитоз медиаторов через мембрану 1-ой клетки. В течение нескольких мкс медиатор достигает постсинаптической мембраны и начинает взаимодействовать с тем её участком, который расположен напротив симпатической щели. Этот участок постсинаптической мембраны, называется субсиноптической мембраной.
Субсиноптическая мембрана существенным образом отличается от остальных участков постсинаптической мембраны. Она не обладает возбудимостью, т.к. не содержит потенциально зависимых каналов. Однако в ней находятся особые гемочувствительные каналы, которые открываются под действием медиаторов. Сенсором медиатора является особые гликопротеидные комплексы, входящие в состав субсиноптической мембраны. Между молекулами медиатора и данными комплексами существует структурное соответствие, обеспечивающее высокую избирательность и их взаимодействие. В субсиноптической мембране находятся ферменты, разрушающие как сами медиаторы так и их комплексы с гликопротеинами. В результате открытия ионных каналов увеличивается чувствительность субсиноптической мембраны. В том случае если открываются натриевые каналы, то возникает градуальная деполяризация субсиноптической мембраны. Если же открываются каливые каналы, то мембрана гиперполяризуется. Так как мембрана не возбудима, то амплитуда де- и гиперполяции определяются числом открытых каналов, т.е. числом медиаторов, переходящих на субсиноптическую мембрану. Деполяризация субсиноптической мембраны может обеспечить возникновения потенциала действия, а, следовательно, и передачу возбуждения второй клетки. Гиперполяция затрудняет синоптическую передачу возбуждения, т.е. тормозит нервный импульс, поэтому все химические синопсы подразделяются на две группы: возбуждение и торможение. В первой группе медиатор возбуждает деполеризацию, во второй – гиперполяцию. При деполяризации на субсиноптической мембране возникает возбуждённый постсиноптический потенциал. Он создаётся за счёт открытия натриевых каналов. Примером такого импульса может быть система, состоящая из ацетилхолина в качестве медиатора и холинорецепторов, входящих в состав субсиноптической мембраны. Адсорбция ацетилхолина на холинорецепторных называют комформационную перестройку молекулы рецептора с образованием внутри него каналов, через который диффундируют внутырь второй клетки ионов натрия. В большинстве синапсов поток ионов через каналы продолжается не более 1мс. В это время комплекс медиатора и рецептора разрушается ферментами. Ионная проницаемость мембраны резко падает и возбуждающий постсиноптический потенциал исчезает. Субсиноптическая мембрана не может генерировать потенциал действия, так как нет потенциал зависимых каналов. Но возбуждающий постсиноптический потенциал декриментно распределяется на соседние области мембраны, обладающий потенциально зависимыми каналами, диполеризация их в том случае, если значение потенциала при достижении канала превышает критический мембранный потенциал. В том числе, возникший потенциал действия распределяется по всем клеткам на все мембраны, достигая области, где существует новая писиноптическая структура и новые медиаторы. Вся последовательность процесса химического синапса занимает значительно больше времени по сравнению с распределением вдоль клетки или при передачи возбуждения электрического энергии синапса. Это время называется синапсной задержкой. Для млекопитающих она составляет 0,3мсек. В случае возникновения на субсиноптической мембране тормозного постсиноптического потенциала, возникший гиперполяризационный сдвиг потенциала дикрементно распределяется вдоль постсиноптисческой мембраны снижая её возбудимость. Одни и те же медиаторы в разных постсинаптических мембранах могут вызвать как возбуждение, так и торможение постсинаптических потенциалов, однако постсинаптические мембраны в 1-й клетки при взаимодействие с данным медиатором дают однозначно одинаковый эффект. С одной клетки могут контактировать присиноптические структуры с разными медиаторами, при этом некоторые из них диполерезуют, другие гиперполерезуют данную клетку. Результирующий эффект определяется алгебраической суммой потенциалов, создающих каждым медиатором. Т.о. клетка представляет с собой интегрированное устройство, число химических синопсов на теле Нерона достигает 3 108
1-пресиноптическая мембрана. 2- синоптическая щель.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|
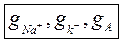

 максимальное удельная электропроводимость мембраны при всех открытых калиевых каналов.
максимальное удельная электропроводимость мембраны при всех открытых калиевых каналов.
 мах. удельная электропроводимость мембран при всех открытых натриевых каналов.
мах. удельная электропроводимость мембран при всех открытых натриевых каналов. - ток смещения
- ток смещения - ёмкость.
- ёмкость.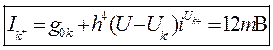 - ток ионов калия
- ток ионов калия - ток ионов натрия
- ток ионов натрия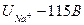






 - константа которая зависит от потенциала на мембране и могут быть определены экспериментально. В зависимости от значений напряжений изменяются величины коэффициентов и соответственно изменяется вклад в общий ток тех или иных ионов, так при достижении потенциала 112мВ происходило открытие каналов Na, при других потенциалах больший вклад давали ионы К.
- константа которая зависит от потенциала на мембране и могут быть определены экспериментально. В зависимости от значений напряжений изменяются величины коэффициентов и соответственно изменяется вклад в общий ток тех или иных ионов, так при достижении потенциала 112мВ происходило открытие каналов Na, при других потенциалах больший вклад давали ионы К.


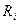 - сопротивление цитоплазмы.
- сопротивление цитоплазмы.

