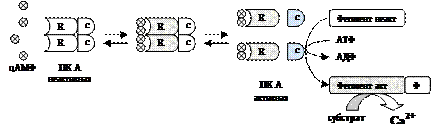|
|
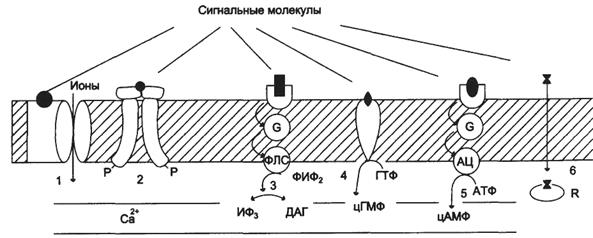
Участие рецепторов в трансмембранной передаче сигнала
1). Рецепторы, связанные с ионными каналами (рецептор ГАМК); 2). Рецепторы, с тирозинкиназной активностью (рецептор инсулина); 3). Рецепторы, активирующие инозитолтрифосфатную систему (α1-адренорецептор - у гепатоцитов); 4). Рецепторы, с гуанилатциклазной активностью (гуанилатциклаза, рецептор ПНФ); 5). Рецепторы, активирующие аденилатциклазную систему (β-адренорецепторы); 6). Рецепторы, связывающие гормон в цитозоле или ядре (рецептор кортизола).
Рецепторы по скорости делятся на: I) быстроотвечающие(в миллисекундах). Это интегральные олигомерные белки с центром связывания сигнальной молекулы и ионным каналом; II) медленноотвечающие(минуты-часы); Медленноотвечающие рецепторы делятся на: 1)каталитические рецепторы, обладающие тирозинкиназной или гуанилатциклазной активностью; Тирозиновые протеинкиназы – это каталитические рецепторы, фосфорилирующие белки по тирозину, они делятся на: а) мембранные, обеспечивают трансмембранную передачу сигнала. Например, рецептор инсулина, гликопротеин, состоит из 2 α и 2 β субъединиц связанных дисульфидными связями. α субъединицы связывают инсулин, а β субъединицы обладают тирозинкиназной активностью. После присоединения гормона к α субъединицам, β субъединицы сначала фосфорилируют друг друга, а затем белок IRS-1, который активирует функциональные ферменты (фосфопротеинфосфатаза). б) цитоплазматические, передают сигнал в ядро. Гуанилатциклазы (ГЦ) – это каталитические рецепторы, превращающие ГТФ в цГМФ. Они находятся на мембране и в цитоплазме. а). Мембранная ГЦ – гликопротеин, активирует гормонами. цГМФ образуемая ГЦ активирует ПК G, ФДЭ, Са2+-АТФазы, закрывает Са2+-каналы и снижает уровень Са2+ в цитоплазме. Разные клетки организма, в зависимости от функций, имеют определенный набор рецепторов. На мембране одной клетки может быть более десятка разных типов рецепторов. б). Цитоплазматическая ГЦ состоит из α и β субъединиц и содержит гем, активируется оксидом азота (NO). Протеинкиназа G (ПК G) ПК G есть только в легких, мозжечке, гладких мышцах и тромбоцитах, участвует в гуанилатциклазной системе. ПК G содержит 2 субъединицы, стимулируется цГМФ, катализирует реакцию фосфорилирования белков-ферментов. 2)рецепторы, действующие на мембранные ферменты через G-белки и активирующие аденилатциклазную и инозитолтрифосфатную систему. По строению эти рецепторы называют серпантинные. У них выделяют внеклеточный домен, внутриклеточный домен и структуру, многократно пронизывающую мембрану. G-белки Универсальные посредники при передаче сигналов от рецепторов к ферментам клеточных мембран, катализирующих образование вторичных посредников гормонального сигнала. G-белки регулируют активность аденилатциклазы, фосфолипазы С, фосфодиэстеразы, Na+- и K+-каналов. G-белки олигомеры, состоят из 3 субъединиц α, β, γ. Есть активаторные Gs-белки, они активируют мембранные ферменты, есть ингибиторные Gi-белки, они ингибируют мембранные ферменты.
Аденилатциклаза (АЦ) Гликопротеин, имеет 8 изоформ, ключевой фермент аденилатциклазной системы, катализирует образование вторичного посредника цАМФ из АТФ. Есть во всех клетках, располагается на внутренней стороне клеточной мембраны. Активность АЦ контролируется: 1) внеклеточными регуляторами - гормонами, эйкозаноидами, биогенными аминами через G-белки; 2) внутриклеточным регулятором Са2+ (4 Са2+-зависимые изоформы АЦ активируются Са2+). Протеинкиназа А (ПК А) ПК А есть во всех клетках. Участвует в аденилатциклазной системе, стимулируется цАМФ. Состоит из 4 субъединиц: 2 регуляторных (R) и 2 каталитических (С). Регуляторные субъединицы имеют по 2 участка связывания цАМФ. Тетрамер не обладает каталитической активностью. Присоединение 4 цАМФ к 2 субъединицам R приводит к изменению их конформации и диссоциации тетрамера. При этом высвобождаются 2 активные каталитические субъединицы С, которые катализируют реакцию фосфорилирования белков-ферментов.
Фосфолипаза С (ФЛ С) Гидролизует фосфоэфирную связь в фосфатидилинозитолах с образованием ДАГ и ИФ3, имеет 10 изоформ. ФЛ С регулируется через G-белки и активируется Са2+. Протеинкиназа С (ПК С) ПК С участвует в инозитолтрифосфатной системе, стимулируется Са2+, ДАГ и фосфатидилсерином. Имеет регуляторный и каталитический домен. ПК С катализирует реакцию фосфорилирования белков-ферментов. Фосфодиэстеразы (ФДЭ) ФДЭ превращает цАМФ и цГМФ в АМФ и ГМФ, инактивируя аденилатциклазную и гуанилатциклазную систему. ФДЭ активируется Са2+, 4Са2+-кальмодулином, цГМФ.

Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|