|
|
Парадигма активности: нейрон, как и индивид, достигает «результат», получая необходимые метаболиты из своей «микросреды»Упомянутые ранее этапы приобретают смысл в том случае, если принять, что процесс, обеспечивающий переход от прек постсинаптическим образованиям, продолжается в непрерывную цепь химических процессов внутри нейрона и, главное, что все межклеточные контакты служат обмену метаболическими субстратами между контактирующими клеточными образованиями. Переход от «поведенческой концепции» к рассмотрению нейрона как организма, получающего необходимые ему метаболиты из окружающей «микросреды», и был тем шагом, который предопределил последующую разработку проблемы в направлении ее системного решения.
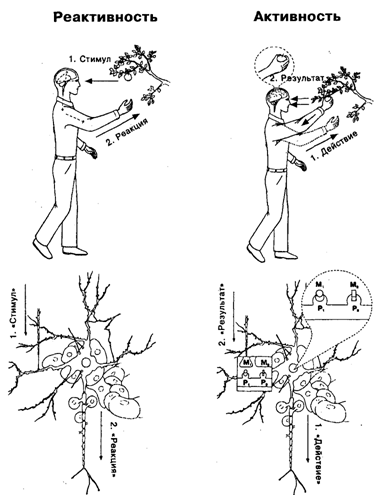
Рис. 14.2. Индивид и нейрон в парадигмах активности и реактивности Цифрами на схеме обозначена последовательность событий. В соответствии с парадигмой реактивности за стимулом (1) следует реакция (2) – поведенческая у человека, импульсная у нейрона. В последнем случае в качестве стимула рассматривается импульсация нейрона, аксон которого (параллельный стрелке с надписью «Стимул») контактирует с дендритом реагирующего нейрона, окруженного глиальными клетками и соприкасающегося с кровеносным сосудом, который расположен над словом «Реакция». Реакция представляет собой импульсные разряды реагирующего нейрона. В соответствии с парадигмой активности действие (1) (поведенческое – у человека, импульсация – у нейрона) завершается достижением результата и его оценкой (2). Пунктирной линией обозначена модель будущего результата: для человека – контакт с объектом-целью, для нейрона – получение соответствующих метаболитов (М1 – от контактирующего нейрона; M2 – от соседней глиальной клетки), которые соединяются с рецепторами нейрона ( P1 , P2) Необходимость дальнейшей разработки определялась тем, что в рамках концепции интегративной деятельности нейрона последовательность событий в принципе оставалась той же, что и в парадигме реактивности. В обоих случаях процесс начинался приходом возбуждения к нейрону и заканчивался генерацией этим нейроном потенциала действия. Разница, которую подчеркивал П.К. Анохин, состояла в том, какими процессами заполнялся интервал между действием медиатора на субсинаптическую мембрану нейрона и генерацией потенциала: химическими преобразованиями внутри нейрона в первом случае и электрической суммацией во втором. Устранение эклектики и приведение представления о детерминации активности нейрона в соответствие с требованиями системной парадигмы было достигнуто отказом от рассмотрения активности нейронов как реакции на синаптический приток и принятием положения о том, что нейрон, как и любая живая клетка, реализует генетическую программу, нуждаясь при этом в метаболитах, поступающих к нему от других клеток [Швырков, 1995]. В связи с этим последовательность событий в деятельности нейрона становится аналогичной той, которая характеризует активный целенаправленный организм, а его импульсация – аналогичной действию индивида (см. рис. 14.2). Активность нейрона, как и поведение организма, является не реакцией, а средством изменения соотношения со средой, «действием», которое обусловливает устранение несоответствия между «потребностями» и микросредой, в частности за счет изменений кровотока, метаболического притока от глиальных клеток и активности других нейронов. Эти изменения, если они соответствуют текущим метаболическим «потребностям» нейрона, приводят к достижению им «результата» (получение набора метаболитов, соединяющихся с его рецепторами; см. далее) и прекращению его импульсной активности. Предполагается, что несоответствие между «потребностями», определяемыми генетически, и реально поступающими метаболитами может иметь место как при генетически обусловленных изменениях метаболизма клетки, так и при изменении притока метаболитов от других клеток. Таким образом, нейрон – не «кодирующий элемент», «проводник» или «сумматор», а организм в организме, обеспечивающий свои «потребности» за счет метаболитов, поступающих от других элементов. Следует подчеркнуть, что для последовательно системного понимания детерминации активности нейрона существенны оба компонента: признание направленности активности нейрона в будущее и ее обусловленности метаболическими «потребностями» нейрона. То, что только первого из них недостаточно, видно на примере интересной концепции гедонистического нейрона, разработанной А. Г. Клопфом [Klopf, 1982]. Утверждая, что целенаправленный мозг состоит из целенаправленных нейронов, А.Г. Клопф отвечает на вопрос о том, в чем нейроны нуждаются и как они это получают в соответствии со следующей логикой. Аристотель рассматривал получение удовольствия как главную цель поведения. Следовательно, организм гедонистичен. Нейрон есть организм. Следовательно, нейрон гедонистичен. «Удовольствие» для нейрона – возбуждение, а «неудовольствие» – торможение. Активация нейрона – «действие», обеспечивающее получение им возбуждения. Нейрон является гетеростатом, т.е. системой, направленной на максимизацию «удовольствия», т.е. возбуждения. Таким образом, отсутствие второго из двух необходимых компонентов ведет к необходимости предположить наличие у нейрона довольно странных и экзотически аргументированных «потребностей», особенно если принять во внимание популярную концепцию «токсического перевозбуждения» (excitotoxic), в рамках которой сильное возбуждение нейронов рассматривается как причина их гибели. В то же время наиболее часто у авторов отсутствует первый из компонентов, что при анализе нейронного обеспечения поведения ведет к рассмотрению сложных метаболических превращений внутри нейрона, главным образом как фактора, обеспечивающего проведение возбуждения и пластичность (модификацию проведения при разных видах научения). При этом сложнейшие механизмы изменения «белкового фенотипа» оказываются направленными, например на изменение чувствительности постсинаптической мембраны к пресинаптическому возбуждению. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|