|
|
ПРОБЛЕМА ВНИМАНИЯ В СИСТЕМНОЙ ПСИХОФИЗИОЛОГИИЭтот парадокс не возникает, если отказаться от рассмотрения поведения как реакции на предъявляемые стимулы. С позиций системной психофизиологии [Швырков, 1995] поведение – это реализация взаимодействующих между собой функциональных систем поведенческих актов разной сложности и с разной историей формирования, представленных нейронами, которые локализованы в самых разных областях нервной системы, и даже рецепторами (см. гл. 14). В рамках этого подхода афферентные и эфферентные влияния рассматриваются не как приход информации в центр и реализация на ее основе моторных программ, а как способ согласования или взаимосодейстия систем (межсистемных отношений). В этом плане демонстративны исследования Ю.И. Александрова [1989]. Исследуя активность ганглиозных клеток сетчатки у кроликов в пищедобывательном поведении, он обнаружил, что эта активность, как и активность центральных нейронов, связана с этапами тщательно контролируемого поведения. Причем активность большинства клеток сохраняет связь с этапами поведения даже в том случае, когда кролик совершает поведение при закрытых специальными светонепроницаемыми колпачками глазах, т.е. при прекращении доступа к ним зрительных раздражителей. Автор связывает этот феномен с включенностью нервных элементов сетчатки глаз в функциональные системы, составляющие индивидуальный опыт животного и обеспечивающие его поведение (см. гл. 14). В других исследованиях, проведенных в рамках системной психофизиологии, было показано, что в конфигурации ССП отражаются не информация, поступившая с периферии, не ее последовательная обработка и не моторная программа, а межсистемные отношения (подробнее см. гл.16). Итак, взаимодействие субъекта с внешним миром возможно только через активность его систем. Набор реализующихся систем определяет специфику поведения, которое может быть выполнено с высокой или низкой степенью эффективности. В этих случаях говорят соответственно о высоком или низком уровне внимания в поведении. Например, скорость и точность отчетных действий в классических задачах выбора рассматриваются как показатели уровня внимания [Блок, 1970]. В задачах выбора испытуемому предъявляют в случайной последовательности через определенный интервал времени разные сигналы (например: а, б, б, а, б ...). В ответ на предъявленный сигнал испытуемый в соответствии с инструкцией должен совершить отчетное действие, например, быстро нажать кнопку «А» при появлении сигнала «а» или быстро нажать кнопку «Б» при появлении сигнала «б».В данной вероятностной ситуации испытуемые сначала пытаются прогнозировать появление следующего сигнала на основании предшествующей последовательности сигналов, что отражается на времени отчетного действия и конфигурации ССП [Remington, 1969; Munson et al., 1984J. Этот феномен известен как эффект последовательности. На основании анализа поведенческих показателей и ССП было выявлено, что в тех случаях, когда наблюдался эффект последовательности, в межсистемные взаимодействия текущего отчетного действия включаются некоторые системы, обеспечивающие предшествующие отчетные действия [Безденежных, 1995; Bezdenezhnykh, 1996]. Однако в процессе тренировки эти «лишние» системы исключаются из обеспечения отчетного действия, время действий сокращается, а количество ошибочных отчетов уменьшается вплоть до полного их исчезновения. Одновременно имеют место устойчивые изменения в ССП: у них изменяются временные и амплитудные характеристики компонентов и они становятся сходными в разных областях мозга. На основании этого можно считать, что в процессе совершенствования деятельности решения задачи выбора формируются и стабилизируются такие межсистемные отношения, которые обеспечивают оптимальную реализацию деятельности. Если же изменения в ССП носят неустойчивый характер, то и показатели внимания неустойчивы [Безденежных, 1995]. Механизм стабилизации межсистемных отношений на таком уровне, который обеспечивает действия в соответствии с инструкцией, недостаточно изучен. Предполагается, что существенную роль здесь играет образ действий, формируемый на основе инструкции [Гордеева, Зинченко, 1982; Безденежных, 1995], и, по мнению С.Л. Рубинштейна [1946], именно во внимании, которое, как уже отмечено, не имеет своего содержания, проявляется взаимосвязь образа и деятельности. В заключение отметим, что с позиций системной психофизиологии внимание рассматривается не как самостоятельный психический процесс, а как отражение таких межсистемных отношений текущей деятельности, которые обеспечивают эффективность этой деятельности. Сходные точки зрения на проблему внимания высказывались в работах ряда исследователей [Найссер, 1981; Величковский, 1982 и др.]. Глава 10 ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС И ОРИЕНТИРОВОЧНО-ИССЛЕДОВАТЕЛЬСКАЯ ДЕЯТЕЛЬНОСТЬ ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС Ориентировочный рефлекс, или рефлекс «Что такое?» был открыт И.П. Павловым. Он описал его как комплекс двигательных реакций, который возникал на неожиданное появление нового стимула. Собака поворачивала голову, глаза, настораживала уши в направлении нового раздражителя. В работах Ю. Конорского [1970] этот комплекс соматических реакций получил название рефлекса прицеливания. Рассматривая ориентировочный рефлекс как реакцию, направленную на восприятие нового стимула, И.П. Павлов обратил основное внимание на его тормозную, прерывающую функцию в отношении текущей условно-рефлекторной деятельности. Во время ориентировочной реакции условный рефлекс либо полностью подавлялся, либо его выполнение ухудшалось. Это свойство ориентировочной реакции нарушать текущую деятельность, в том числе и процесс обучения (выработку условного рефлекса), привело И.П. Павлова к мысли о необходимости построить исследовательский корпус в виде «Башни молчания». Это позволило бы исключить тормозные эффекты ориентировочного рефлекса (ОР) на формирование и исполнение условных рефлексов. Хотя вначале ОР и представлялся И. П. Павлову и его коллегам преимущественно как внешний тормоз условного рефлекса, они описали многие его особенности, которые выходили за пределы тормозной функции. Так был открыт растормаживающий эффект ориентировочной реакции. Он состоял в том, что ранее выработанные, а затем угашенные условные связи под влиянием ОР вновь восстанавливались. Сегодня мы интерпретировали бы эти данные как способность восстанавливать подавленные связи в процедурной долговременной памяти. В лаборатории И.П. Павлова также были получены первые факты о влиянии ОР на формирование условного рефлекса. Во время процедуры выработки условной связи, а также при формировании ассоциации между двумя индифферентными раздражителями было отмечено усиление или появление ориентировочных реакций. В опытах И.О. Нарбутович и Н.А. Подкопаева [1936] с выработкой у собаки ассоциации в виде двух последовательно следующих индифферентных стимулов (звук – свет или вертушка – прерывистый шум) было показано, что при отсутствии на них ориентировочных реакций ее формирование идет плохо. В этих же опытах по ОР судили о сформированности ассоциации. Когда собака вырабатывала связь между звуком и светом, можно было видеть, что она поворачивает голову в сторону света еще до его предъявления. Однако на том этапе изучения ориентировочного рефлекса еще не сложилось полного представления как о его взаимоотношениях с условным рефлексом, так и о физиологических механизмах и структуре самого ориентировочного рефлекса. П. К. Анохин выделил три формы взаимоотношений ориентировочного рефлекса с текущей деятельностью. К первой он относит «конфликтные отношения», когда ориентировочная активность оказывает тормозный эффект. Эти отношения детально были изучены в лаборатории И.П. Павлова. Вторая форма отражает ассимиляцию ориентировочной реакции оминирующей деятельностью. В этом случае ОР не тормозит, а усиливает текущую деятельность. Например, в ситуации оборонительной доминанты появление нового стимула вызывает не ориентировочный, а оборонительный рефлекс. К третьей форме отношений относится трансформация ОР в поведенческую реакцию, связанную со скрытой, внешне непроявляющейся доминантой. Анализируя эти данные, П.К. Анохин [1968] пришел к выводу том, что эффект ОР существенно зависит от предыдущего опыта животного и от ранее сформировавшихся и зафиксированных в памяти доминант. Открытие в 1929 г. Г. Бергером альфа-ритма человека и последующее совершенствование техники записи волновой активности мозга позволили связать ОР с депрессией альфа-ритма. Важное значение для изучения ОР имело открытие Г. Моруцци и X. Мэгуном (1949) ретикулярной активирующей системы мозга, позволившее связать блокаду альфа-ритма с реакцией активации, регулируемой этой системой. Наиболее всесторонне ориентировочный рефлекс был исследован в работах Е.Н. Соколова и его коллег. Полиграфическая регистрация электроэнцефалографических, вегетативных и двигательных реакций животных и человека в условиях многократного повторения индифферентных стимулов привела к представлению об ориентировочном рефлексе как о многокомпонентной реакции активации. Е.Н. Соколов [I960], а затем Р. Линн [Lynn, 1966] описали систему компонентов ОР. На уровне неокор-текса ОР представлен ЭЭГ-реакцией активации (блокадой альфа-ритма, усилением бетаи гаммаколебаний), коррелирующей с увеличением возбудимости, реактивности и лабильности у нейронов коры. К вегетативным компонентам ОР относят расширение зрачков, увеличение кожной проводимости, снижение ЧСС, изменение дыхания, увеличение дыхательной аритмии в частотном спектре ритмограммы сердца, расширение сосудов головы и сужение сосудов рук. В моторной сфере ориентировочный рефлекс представлен поворотом головы, движением глаз, настораживанием ушей в направлении нового стимула и увеличением мышечного тонуса, обеспечивающего повышенную готовность к моторным реакциям. Под влиянием ОР сенсорная чувствительность анализаторов, измеряемая по субъективным реакциям, увеличивается, что получило название сенсорного компонента ориентировочного рефлекса. Характер электроэнцефалографических, вегетативных и моторных реакций во время ОР позволяет понять суть рефлекса «Что такое?». Ориентировочный рефлекс обеспечивает лучшее восприятие вызвавшего его стимула и готовность быстро реагировать, если того потребует ситуация. Ради этого прерывается текущая деятельность и ресурсы внимания обращаются на новый стимул. Отличительной особенностью ОР является его привыкание или угашение, развивающееся при повторении индифферентного стимула. Е.Н. Соколов [1958] выделил две формы ориентировочной реакции: генерализованную и локальную, которые выражают разные этапы процесса угашения ОР. В зависимости от длительности ОР различают тонический и фазический ориентировочный рефлекс. Первоначально новый стимул вызывает генерализованный ОР, связанный с возбуждением ретикулярной формации ствола мозга. Он характеризуется ЭЭГ-активацией, которая охватывает всю кору в течение достаточно длительного времени (тоническая форма ОР, часто возникающая параллельно с генерализованным ОР; см. гл. 8). Ее легко увидеть у субъекта, находящегося в дремотном состоянии. Тогда говорят о реакции пробуждения (arousal reaction). Подобную реакцию также наблюдали Шарплесс и Джаспер [Sharpless, Jasper, 1956] у кошек и назвали ее тонической реакцией пробуждения. Генерализованный ориентировочный рефлекс быстро угасает, обычно после 10–15 применений индифферентного стимула. Это позволяет выявить локальный ориентировочный рефлекс, который более устойчив к угашению и требует 30 и более применений стимула. Его генерацию связывают с активацией неспецифического таламуса. При локальном ОР реакция ЭЭГ-активации сохраняется в сенсорной коре, соответствующей модальности повторяемого раздражителя. Так, свет продолжает вызывать блокаду альфа-ритма, но только в зрительной коре, звук – блокаду тау-ритма, регистрируемого от височной области, тактильное и проприоцептивное раздражение при пассивном подъеме пальца руки вызывает депрессию мю-ритма (роландического ритма) локально в сенсомоторной коре. Длительность реакции сокращается, она становится фазической, появляясь только на включение и выключение стимула (фазическая реакция пробуждения по Шарплису и Джасперу). Продолжая повторять стимул, можно добиться полного угашения ОР. Ориентировочный рефлекс с полным набором компонентов возникает не у всех испытуемых, что выражает их индивидуальные различия. Изучение компонентного состава ОР на звуковой стимул (1000 Гц, 50 дБ) у 100 испытуемых показало, что блокада альфа-ритма в зрительной коре наблюдается у 85 человек, блокада роландического – у 53, появление КГР – у 95, движение глаз возникало у 42 испытуемых, изменение частоты дыхания – у 63, а изменение мышечного тонуса пальца наблюдалось только у одного испытуемого. Все компоненты ОР (за исключением реакции мышц) были представлены только у 11 испытуемых из 100 (Воронин, Соколов, 1960). Хотя ориентировочный рефлекс, и относится к безусловным рефлексам, он обнаруживает способность к угашению (так как для него специфическим стимулом является «новизна», а специфической реакцией – набор компонентов ориентировочного рефлекса). Это сближает его с условным рефлексом, который сам по себе является продуктом обучения. Именно поэтому угашение ОР рассматривается как особая форма обучения, получившего название негативного обучения, которое ведет к потере способности у данного стимула вызывать ориентировочные реакции. Угашение ОР характеризуется высокой избирательностью к физическим характеристикам стимула. Это проявляется в том, что даже небольшое изменение любого из параметров повторяющегося стимула ведет к восстановлению ОР. Стимулы, сильно отличающиеся от угашаемого, вызывают более сильные ориентировочные реакции, а стимулы, близкие ему – более слабые. Величина ОР увеличивается пропорционально количеству одновременно измененных параметров стимула. Для объяснения этого явления Е.Н. Соколов предположил, что в процессе повторения индифферентного стимула без специального подкрепления в мозге формируется его нейрональная модель (см. также гл. 9). Мозг замечает малейшее отклонение от параметров повторяющегося стимула благодаря сравнению с его моделью, хранящейся в памяти, будь то изменение интенсивности, длительности, цвета, формы, частоты и т.д. В нервной модели запечатлеваются не только элементарные, но и сложные признаки, например комплексы раздражителей – совместное или последовательное применение нескольких раздражителей. Исключение одного из элементов комплекса или изменение их порядка вызывают ориентировочные реакции. Нервная модель стимула многомерна, она фиксирует одновременно все его параметры, что доказывается восстановлением ОР при изменении любого физического свойства повторяющегося стимула даже в случае уменьшения интенсивности или укорочения его длительности. Новизна как специфический тригер безусловного ОР определяется сигналом рассогласования, генерируемым при несовпадении нервной модели стимула с возбуждением от предъявляемого раздражителя. Схема взаимодействия основных функциональных блоков с нервной моделью, по Е.Н. Соколову, объясняющая угашение и восстановление ОР, включает воспринимающее устройство, исполнительное устройство ОР, блок с моделью стимула, компаратор (устройство, в котором осуществляется операция сравнения стимула с сигналом, поступающим от блока с моделью), блок активирующей системы, определяющий уровень активности исполнительного устройства ориентировочного рефлекса. По мере формирования модели она усиливает свое тормозное влияние на неспецифическую систему мозга. Торможение носит избирательный характер, оно ухудшает восприятие только повторяющегося стимула. В случае несовпадения стимула с моделью в компараторе возникает сигнал рассогласования, который активирует исполнительное устройство ОР. Следует отметить, что некоторые исследователи увеличивают количество факторов, вызывающих ОР. Класс явлений, названных Д. Берлайном «сличительными признаками», которые стимулируют ориентировочно-исследовательское поведение, включает, помимо новизны, удивление, неопределенность, двусмысленность и сложность. Однако, по мнению Р. Линна [Lynn, 1966], они могут быть интерпретированы как разные формы новизны. Так, удивление связано с появлением чего-то нового в последовательности стимулов, а сложность, необычность и двусмысленность представляют новизну в паттерне стимулов. Изучение нейронных коррелятов ориентировочного рефлекса выявило существование особого класса нейронов – нейронов новизны, которые были обнаружены в разных структурах мозга. Впервые нейроны новизны были найдены в зрительной покрышке лягушки Дж. Летвиным и др. [1964]. Эти клетки переставали отвечать на повторяющееся движение зрительного объекта, совершаемое в одном и том же направлении. Изменение траектории движения восстанавливало ответ нейрона в виде спайковых разрядов. Нейроны новизны были эфферентными, а их длинные аксоны оканчивались в сетчатке. Эти нейроны сходны с «нейронами внимания», выделенными Д. Хьюбелом и др. (Hubel et al., 1959) в слуховой коре кошки. Нейроны со сходными характеристиками были также найдены в ретикулярной формации ствола мозга кошки (Huttenlocher, 1961; Lindsley et al., 1973). Изучение неспецифического таламуса кролика показало существование особой формы нейронной реакции активации, отличной от простого учащения спайковых разрядов. Новые стимулы вызывали десинхронизацию спайковой активности нейрона, которая состояла из разрушения его пачечной активности, коррелирующей с волнами ЭЭГ, и замены ее на рандоминизированные одиночные спайки, появление которых совпадало с блокадой альфа-подобной активности [Данилова, 1970, 1985]. Реакция десинхронизации нейронов таламуса демонстрирует все свойства ориентировочной реакции. Вначале формирование нервной модели стимула Е.Н. Соколов связывал с функцией корковых нейронов. Однако после открытия О.С. Виноградовой в поле САЗ гиппокампа двух групп нейронов, реакции которых обнаружили характеристики ОР (В-нейроны, реагирующие возбуждением, и Т-нейроны, отвечающие на новый стимул торможением), он связал нервную модель стимула с нейронами гиппокампа. Е.Н.Соколов [1995] рассматривает эти две группы нейронов гиппокампа как «нейроны новизны» (В-нейроны) и «нейроны тождества» (Т-нейроны). Их совместное действие позволяет подчеркивать новые раздражители и ослаблять действие привычных стимулов за счет противоположного влияния на активирующую и инактивирующую части модулирующей системы мозга. Формирование нервной модели определяется тем, что множество нейронов-детекторов конвергируют на нейроны поля САЗ гиппокампа (нейроны новизны и тождества). Они создают на каждом из них универсальное рецептивное поле, охватывающее всю рецепторную поверхность животного [Соколов, 1981]. При действии стимула происходит селективное включение детекторов. С повторением раздражителя возбуждение детекторов сохраняется прежним. Однако меняются их синаптические контакты на нейронах гиппокампа. Веса задействованных пластических синапсов на гиппокампальных нейронах уменьшаются пропорционально силе приходящего возбуждения. В результате возбудительный ответ нейрона новизны на повторяющийся стимул подавляется и от него регистрируется лишь фоновая активность. С повторением стимула параллельно меняется и реакция нейронов тождества, с которыми детекторы связаны не возбуждающими, а тормозными пластическими синапсами. Их тормозная реакция на новый стимул постепенно уменьшается, и нейрон сохраняет свою фоновую активность. Е.Н. Соколов предположил, что реципрокные сигналы от нейронов новизны и тождества противоположно воздействуют на активирующую и синхронизирующую системы мозга. Новый раздражитель вызывает реакцию активации, так как возбуждает активирующую и тормозит синхронизирующую (инактивирующую) системы мозга. Привычный стимул перестает возбуждать активирующую неспецифическую систему и при этом через нейроны тождества стимулирует неспецифические тормозные влияния. Таким образом, нервная модель стимула представлена на нейронах новизны и тождества матрицами потенциированных синапсов, связанных со свойствами раздражителя и отражающими его конфигурацию. Новый стимул активирует новые синапсы нейронов новизны и тождества, еще не подвергнувшихся пластическим перестройкам. Это определяет появление ориентировочного рефлекса за счет возбуждения активирующей и подавления инактивирующей систем мозга. Величина ОР увеличивается пропорционально величине различия (рассогласования) между новым стимулом и сформированной нервной моделью [Соколов, 1995]. Дальнейший прогресс в понимании механизма ОР связан с применением компьютерной техники усреднения при изучении потенциалов мозга, связанных с событиями (ССП; см. гл. 16). Значительный вклад в идентификацию и классификацию различных компонентов ССП и в поиски мозговых аналогов сигналов рассогласования внесли работы Р. Наатанена и его коллег. Им выявлена специфическая мозговая волна, названная негативностью рассогласования (HP). Она получена в опытах, в которых в случайном порядке и с короткими межстимульными интервалами, (не более 10–14 с) следовали стандартный (часто повторяющийся, 80–90%) и девиантные (несколько отклоняющиеся от стандарта и редко предъявляемые, 20–10%) стимулы. При этом внимание испытуемого было направлено на чтение интересной книги. Р. Наатанен для извлечения HP предложил процедуру вычитания ССП на стандартный стимул из ССП на девиантный стимул. Им представлено большое количество экспериментальных доказательств связи HP с операцией сличения действующего стимула с короткоживущим (не более 20 с) следом стандартного стимула в памяти. Чем сильнее они отличаются, тем больше амплитуда и короче латенция HP [Наатанен, 1997]. Исследователями выявлено два компонента HP: модально-специфический и фронтальный. Определяя локусы генерации модально-специфической HP для звуковых стимулов с помощью МЭГ, они нашли их в специфической слуховой коре, где, как предполагает Р. Наатанен, хранятся короткоживущие сенсорные следы. Для разных параметров звуковых стимулов (частоты, интенсивности и пространственного расположения) обнаружены разные HP с несовпадающими локусами их генерации. Предполагают, что сенсорно-специфическая HP трансформируется во фронтальную HP. Процесс сличения стимула с короткоживущим следом памяти, который представлен в HP, протекает автоматически и не требует сознания. В частности, это подтверждается тем, что HP может возникать и на подпороговые девиации. При этом чем больше амплитуда подпороговой HP, тем ниже субъективный порог различения испытуемого. HP рассматривается как начальная фаза обработки информации о сигнале, за которой может следовать фаза непроизвольного внимания (безусловный ориентировочный рефлекс) к девиантному стимулу. Р. Наатанен связывает его с волной Н2б. HP как выражение сигнала рассогласования отличается от классического ОР, который, как уже упоминалось, также возникает в результате рассогласования нервной модели стимула и наличного возбуждения от раздражителя. Однако в последнем случае время жизни нервной модели значительно больше, что позволяет при угашении ОР и формировании нервной модели использовать длительные межстимульные интервалы порядка нескольких десятков секунд. Сравнивая HP с классическим ОР, Р.Наатанен [1997] отмечает, что процесс генерации сенсорно-специфической HP имеет очевидное сходство с процессом рассогласования, постулируемым Е.Н. Соколовым для возникновения ОР, и в то же время между ними имеется существенное различие. HP – первое событие, возникающее в активности мозга, которое направлено на инициацию ОР в ответ на физическое изменение стимула, когда стимулы предъявляются с достаточно короткими межстимульными интервалами, чтобы вызвать HP. Появление ОР в начале стимульной последовательности и в ответ на изменение стимула при длительных межстимульных интервалах имеет другой механизм, отличный от вызова ОР на основе HP. Другой сигнал рассогласования, который более тесно связан с классическим ОР, был выявлен М. Фербатеном и др. [Verbaten et al., 1986]. Для этого был использован не метод усреднения ССП, а процедура удаления шумового компонента, что позволило исследовать динамику одиночных ССП на стимулы, предъявляемые с большими интервалами. Исследователи нашли, что при угашении ОР (его компонента в виде увеличения кожной проводимости пальцев) параллельно идет процесс уменьшения неспецифической части волны Н1. Новый стимул восстанавливает неспецифический компонент H1 и реакцию повышения кожной проводимости. Выделенная часть компонента Н1 демонстрирует неспецифические свойства ОР и зависимость от долгоживущего следа памяти (десятки секунд). Таким образом, сигналы рассогласования, участвовавшие в генерации непроизвольного ОР, возникают по крайней мере на двух разных типах следовых процессов: короткоживущей и долгоживущей моделях стимула. Следует отметить, что теория ориентировочного рефлекса Е.Н. Соколова, несмотря на ее большую популярность, все же подвергалась критике. Так, отмечалось, что с ее позиций трудно объяснить появление ОР в случаях когнитивного рассогласования, т.е. когда не подтверждается ожидание относительно последовательности стимулов [Underwood, Moray, 1971]. Например, ориентировочная реакция возникает на цифру «9» в последовательности 1, 2, 3, 4, 5, 6, 7, 9. Другое направление критики связано с утверждением о том, что теория ОР Е.Н. Соколова не объясняет появления ОР на значимые стимулы [Bernstein, 1979; Maltzman, 1979]. Сторонники концепции значимости считают, что решающее значение для появления ОР имеют корковые процессы интерпретации и усиления. С этих позиций теория ОР, возникающего на любое физическое изменение стимула, рассматривается как не оринтированная на реальное поведение и деятельность [Bernstein, 1969]. Однако участники полемики относительно значимости ОР не учитывают, что наряду с безусловным ОР Е.Н. Соколов [1958] вводит понятие условного ориентировочного рефлекса, возникающего на значимый сигнальный стимул. Он показал, что придание раздражителю сигнального значения (обычно с помощью инструкции или речевого подкрепления, побуждающих человека отвечать определенной реакцией на определенный стимул) восстанавливает и усиливает ОР на условный сигнал. Эффект усиления ОР сохраняется до тех пор, пока условный рефлекс не достигнет стадии автоматизации. На рис. 10.1 представлено усиление КГР, измеряемого как падение сопротивления кожи, по Фере, при придании звуковому стимулу сигнального значения. По инструкции испытуемый должен был сжимать руку в кулак при предъявлении звука после предварительного угашения на него безусловного ОР. Выработку условного ориентировочного рефлекса можно увидеть при формировании ассоциации: звук 70 дБ – звук 90 дБ (рис. 10.2). После угашения КГР на индифферентный звук 70 дБ присоединение к нему второго стимула – 90 дБ восстанавливает реакцию, которая усиливается по мере сочетаний стимулов. Постепенно КГР-реакция на комплекс уменьшается и исчезает. Однако изолированное применение стимулов вызывает усиленный ОР, выражающий рассогласование стимула со сформированной в памяти моделью комплекса. Сравнивая форму ССП на редко предъявляемые девиантные стимулы, когда они были релевантными и испытуемый должен был на них как-то реагировать и когда те же стимулы им игнорировались (испытуемый читал книгу), Р. Наатанен выделил среди компонентов ССП особый потенциал, который коррелировал с произвольным вниманием. Эта волна получила название процессной негативности (ПН). Под влиянием инструкции быть внимательным к особым стимулам, ПН усиливается на все стимулы, но особенно на релевантные. Чем короче межстимульный интервал, тем раньше возникает ПН (в среднем ее ЛП около 150 мс, но она может появляться и через 50 мс).
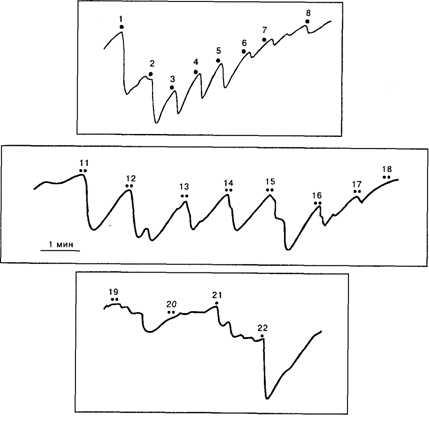
Рис.10.1 . Усиление КГР – компонента ориентировочного рефлекса на звук, на который испытуемый должен был реагировать движением руки (10 – 24): 9 – инструкция; 1–8– индифферентные звуковые стимулы до инструкции. Видно, что усиление фазических КГР-реакций сопровождается тоническим снижением сопротивления кожи, которое сохраняется до 24-го применения стимула. По оси ординат – сопротивление в килоомах [Соколов, 1958]
Рис.10.2.Выработка условного ориентировочного рефлекса при формировании ассоциации двух звуковых стимулов: 1-8 - звук 1000 Гц, 70 дБ, 5 с; 11–20 - комплекс: 70 дБ - 90 дБ; 21 - 70 дБ, 22– 90 дБ. По оси ординат – сопротивление в килоомах [Соколов, 1958] ПН не тождественна Н 1, хотя и совпадает с ней по латенции. Критическим признаком, отличающим Н 1 от ПН, является его инверсия при смещении активного электрода по слуховой коре относительно мостадоидов (сосцевидных отростков). Объясняя ПН, Р. Наатанен [1997] вводит понятие «следа селективного внимания», который связан с памятью. Он полагает, что произвольное внимание актуализирует в сенсорной памяти (по-видимому, во вторичной ассоциативной коре) след релевантного стимула. «След внимания» понимается им как механизм опознания стимула, так как входной сигнал всякий раз сличается со следом релевантного стимула, который непрерывно поддерживает в активном состоянии произвольное внимание. Чем ближе стимул к релевантному, тем больше ПН. Р. Наатанен выделяет раннюю ПН с локусом в ассоциативной коре, выражающей завершение процесса сличения, и позднюю ПН с фронтальной локализацией. Последняя отражает произвольное усилие, направленное на дальнейшую обработку стимула, включающее произвольный ОР на значимый стимул. Суммируя результаты собственных исследований с данными Р. Наатанена и его коллег, Е.Н. Соколов [1997] выделяет два типа энграмм, с которыми связано возникновение непроизвольного и произвольного ОР. Когда единицы кратковременной памяти подключены к нейронам новизны и тождества прямо в виде потенциированных синапсов от детекторов-признаков, сигнал рассогласования, возникающий при несовпадении стимула с энграммой, определяет генерацию непроизвольного ОР. Его сигнал рассогласования представлен в HP и неспецифическом компоненте Н 1. Если же к нейронам новизны и тождества подключены единицы долговременной памяти и они обеспечивают произвольное удержание шаблона, то совпадение стимула с шаблоном порождает сигнал согласования или совпадения. В терминах нейронных популяций он представлен ПН. Сигнал совпадения достигает нейронов активирующей системы и вызывает условный ОР, направленный на значимый стимул. В случае несовпадения стимула с шаблоном сигнал совпадения и ПН не возникают и ОР на значимый стимул не появляется. При этом стимулы, отличающиеся от релевантного, вызывают безусловный ОР, непроизвольное внимание, которое представлено негативностью рассогласования и неспецифическим компонентом Н1. 
Не нашли, что искали? Воспользуйтесь поиском по сайту: ©2015 - 2026 stydopedia.ru Все материалы защищены законодательством РФ.
|